

Myosin IIB assembly state determines its mechanosensitive dynamics
- PMID: 30655296
- PMCID: PMC6400566
- DOI: 10.1083/jcb.201806058
Myosin IIB assembly state determines its mechanosensitive dynamics
Abstract
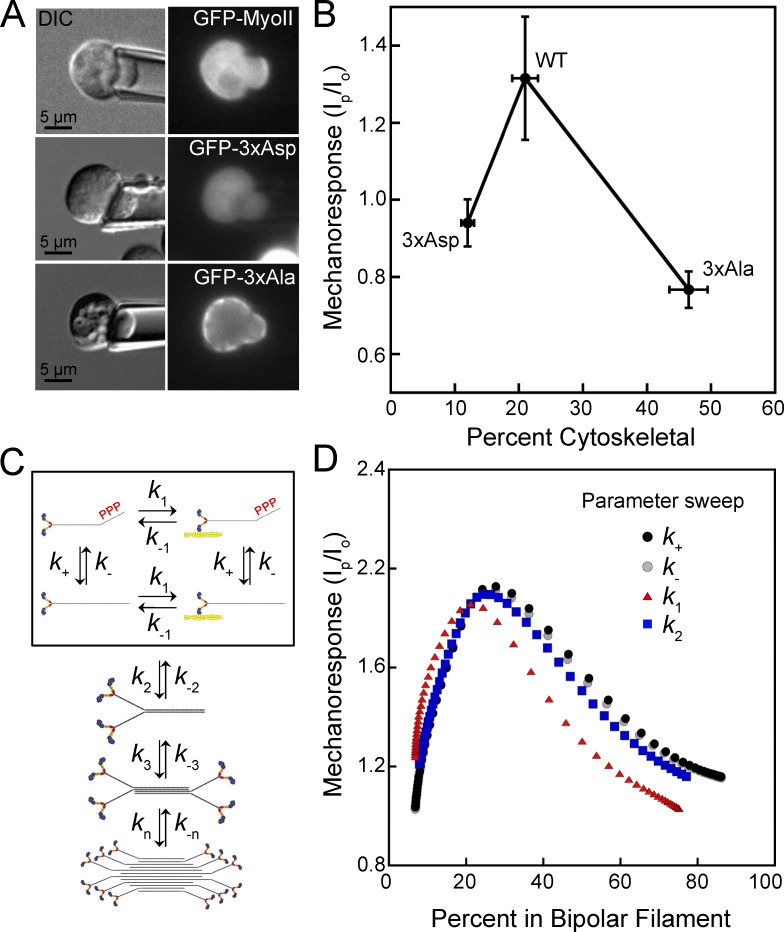
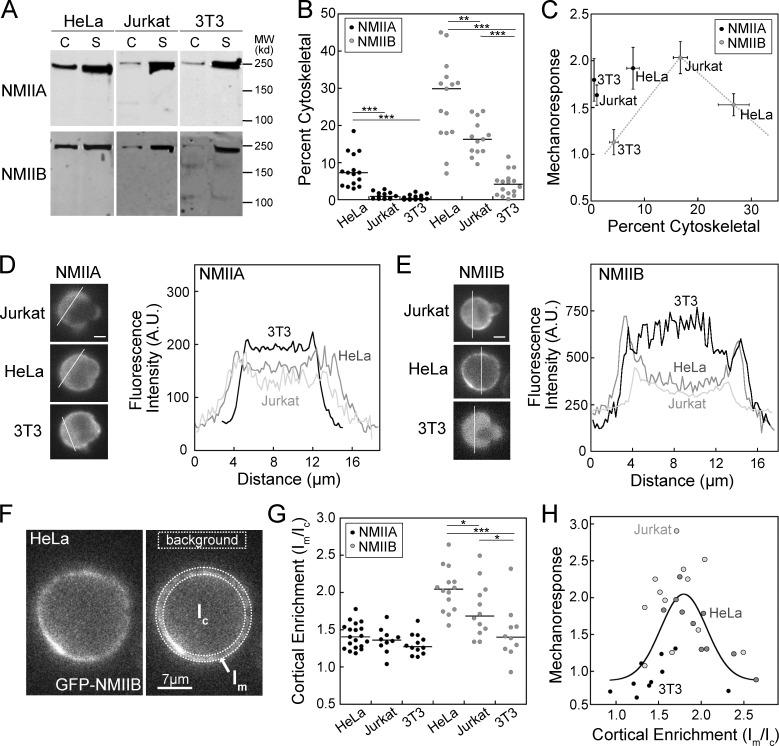
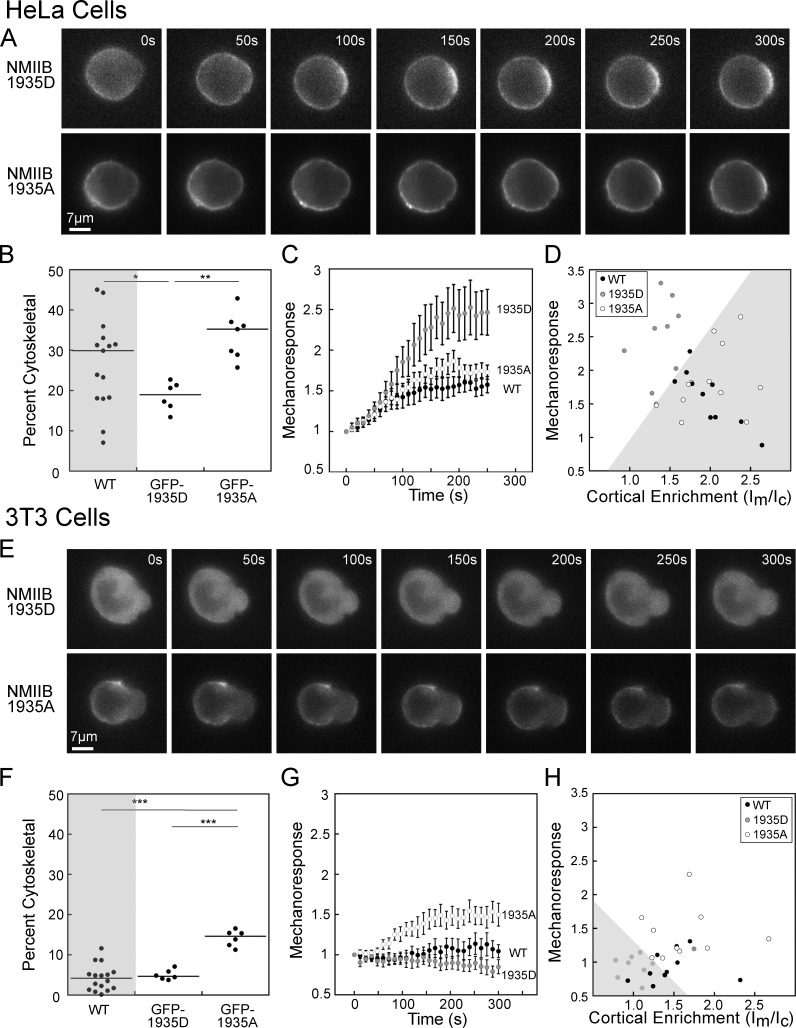
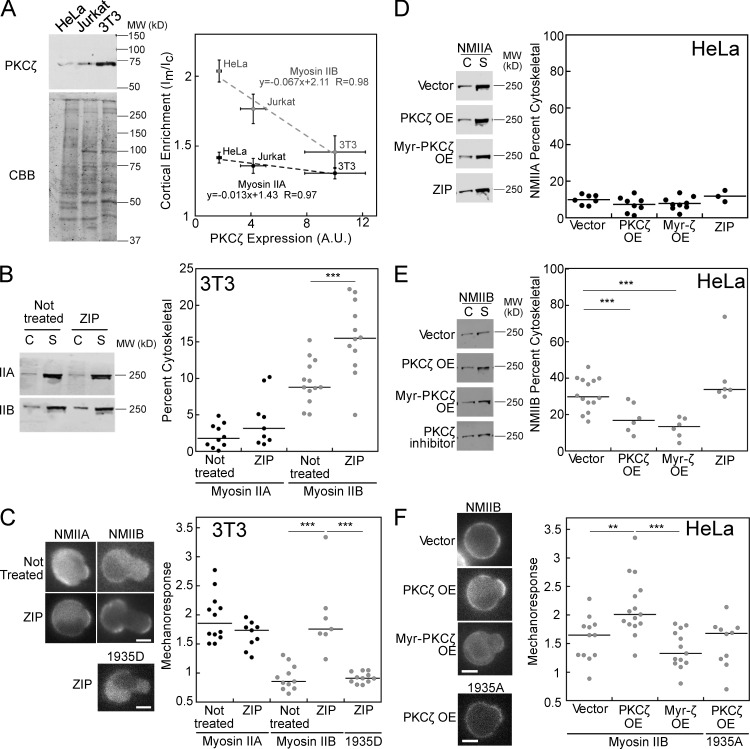
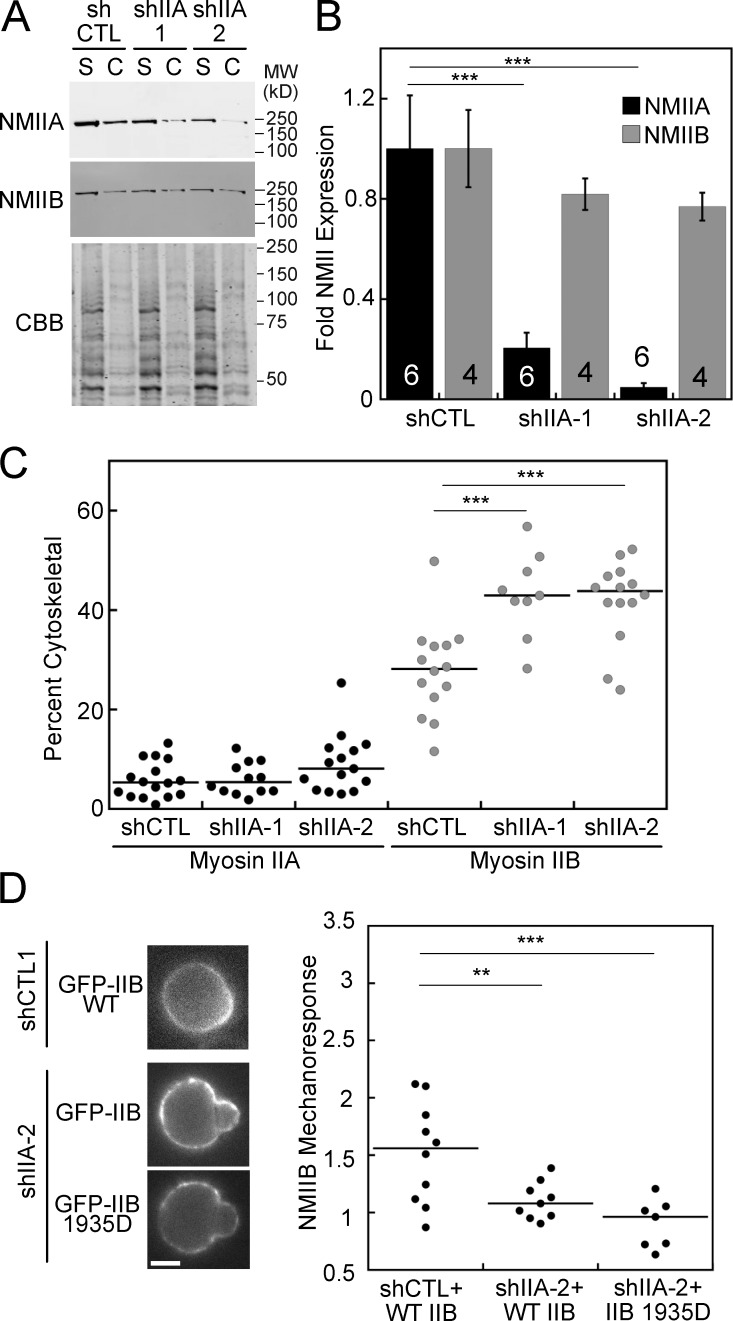
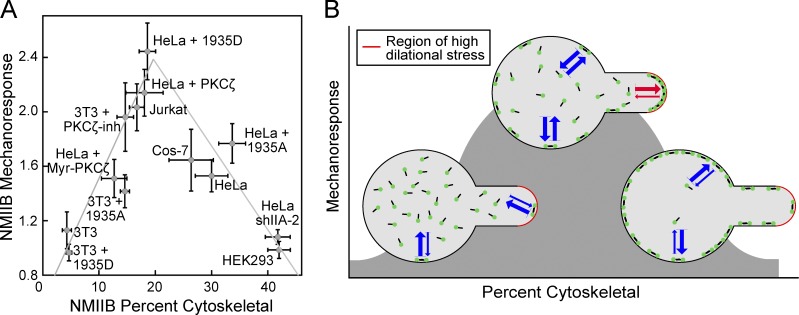
Dynamical cell shape changes require a highly sensitive cellular system that can respond to chemical and mechanical inputs. Myosin IIs are key players in the cell's ability to react to mechanical inputs, demonstrating an ability to accumulate in response to applied stress. Here, we show that inputs that influence the ability of myosin II to assemble into filaments impact the ability of myosin to respond to stress in a predictable manner. Using mathematical modeling for Dictyostelium myosin II, we predict that myosin II mechanoresponsiveness will be biphasic with an optimum established by the percentage of myosin II assembled into bipolar filaments. In HeLa and NIH 3T3 cells, heavy chain phosphorylation of NMIIB by PKCζ, as well as expression of NMIIA, can control the ability of NMIIB to mechanorespond by influencing its assembly state. These data demonstrate that multiple inputs to the myosin II assembly state integrate at the level of myosin II to govern the cellular response to mechanical inputs.
© 2019 Schiffhauer et al.
Figures
Similar articles
-
Endogenous species of mammalian nonmuscle myosin IIA and IIB include activated monomers and heteropolymers.Curr Biol. 2014 Sep 8;24(17):1958-68. doi: 10.1016/j.cub.2014.07.070. Epub 2014 Aug 14. Curr Biol. 2014. PMID: 25131674 Free PMC article.
-
Differential contributions of nonmuscle myosin IIA and IIB to cytokinesis in human immortalized fibroblasts.Exp Cell Res. 2019 Mar 1;376(1):67-76. doi: 10.1016/j.yexcr.2019.01.020. Epub 2019 Jan 31. Exp Cell Res. 2019. PMID: 30711568
-
Mosaic loss of non-muscle myosin IIA and IIB is sufficient to induce mammary epithelial proliferation.J Cell Sci. 2017 Oct 1;130(19):3213-3221. doi: 10.1242/jcs.208546. Epub 2017 Aug 18. J Cell Sci. 2017. PMID: 28821574 Free PMC article.
-
The regulation of myosin II in Dictyostelium.Eur J Cell Biol. 2006 Sep;85(9-10):969-79. doi: 10.1016/j.ejcb.2006.04.004. Epub 2006 Jun 30. Eur J Cell Biol. 2006. PMID: 16814425 Review.
-
Signaling pathways regulating Dictyostelium myosin II.J Muscle Res Cell Motil. 2002;23(7-8):703-18. doi: 10.1023/a:1024467426244. J Muscle Res Cell Motil. 2002. PMID: 12952069 Review.
Cited by
-
Structure, regulation, and mechanisms of nonmuscle myosin-2.Cell Mol Life Sci. 2024 Jun 15;81(1):263. doi: 10.1007/s00018-024-05264-6. Cell Mol Life Sci. 2024. PMID: 38878079 Free PMC article. Review.
-
Particle-based model of mechanosensory contractility kit assembly.Biophys J. 2022 Dec 6;121(23):4600-4614. doi: 10.1016/j.bpj.2022.10.031. Epub 2022 Oct 22. Biophys J. 2022. PMID: 36273263 Free PMC article.
-
Molecular Control of Non-Muscle Myosin II-A Aggregation and Intracellular Dynamics by motor- or tail-specific MYH9 Mutations.bioRxiv [Preprint]. 2025 May 21:2025.05.20.654665. doi: 10.1101/2025.05.20.654665. bioRxiv. 2025. PMID: 40475454 Free PMC article. Preprint.
-
The lectin Discoidin I acts in the cytoplasm to help assemble the contractile machinery.J Cell Biol. 2022 Nov 7;221(11):e202202063. doi: 10.1083/jcb.202202063. Epub 2022 Sep 27. J Cell Biol. 2022. PMID: 36165849 Free PMC article.
-
MYH10 Governs Adipocyte Function and Adipogenesis through Its Interaction with GLUT4.Int J Mol Sci. 2022 Feb 21;23(4):2367. doi: 10.3390/ijms23042367. Int J Mol Sci. 2022. PMID: 35216482 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
