

A De Novo Shape Motif Discovery Algorithm Reveals Preferences of Transcription Factors for DNA Shape Beyond Sequence Motifs
- PMID: 30660610
- PMCID: PMC6368855
- DOI: 10.1016/j.cels.2018.12.001
A De Novo Shape Motif Discovery Algorithm Reveals Preferences of Transcription Factors for DNA Shape Beyond Sequence Motifs
Abstract
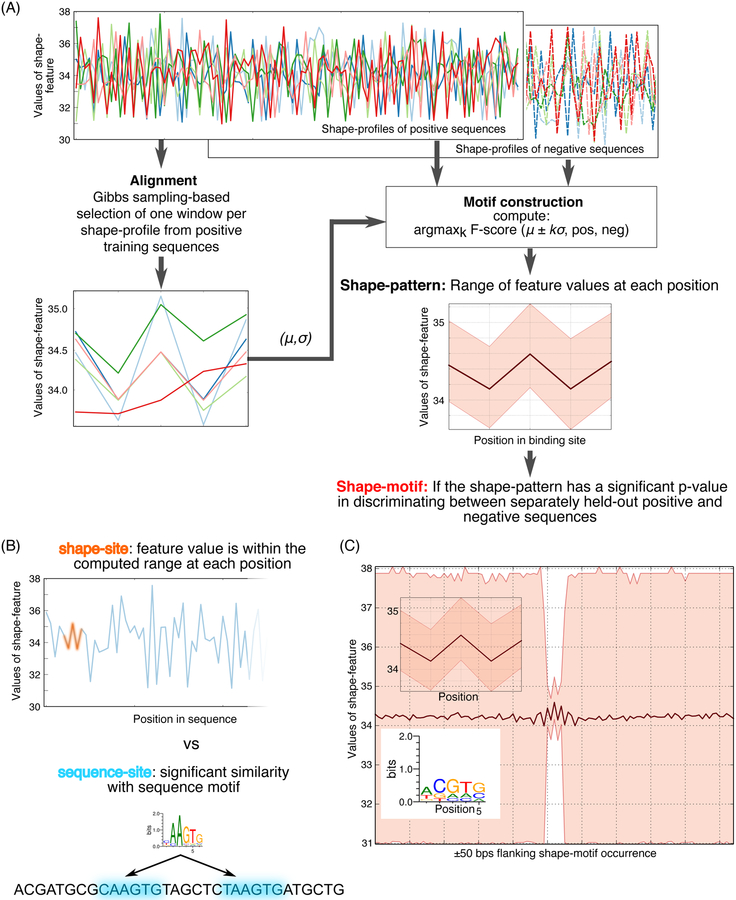
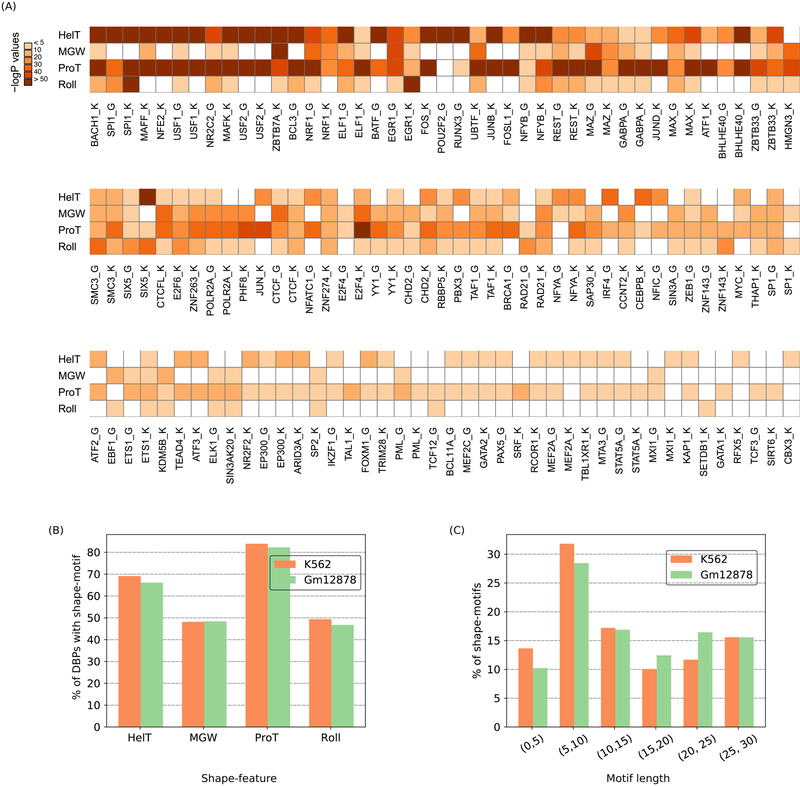
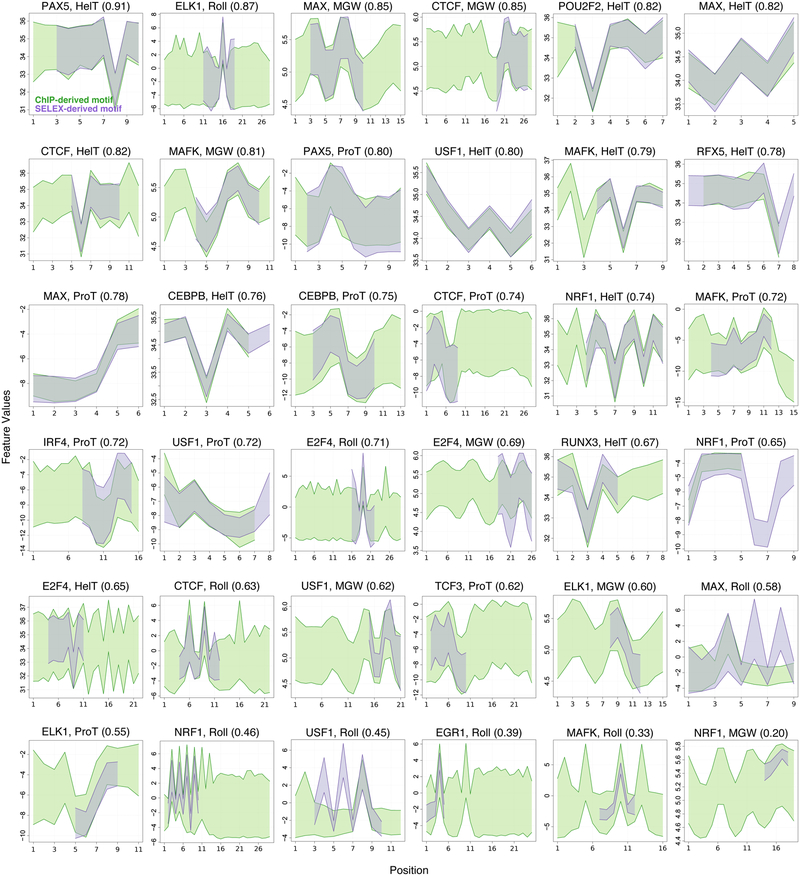
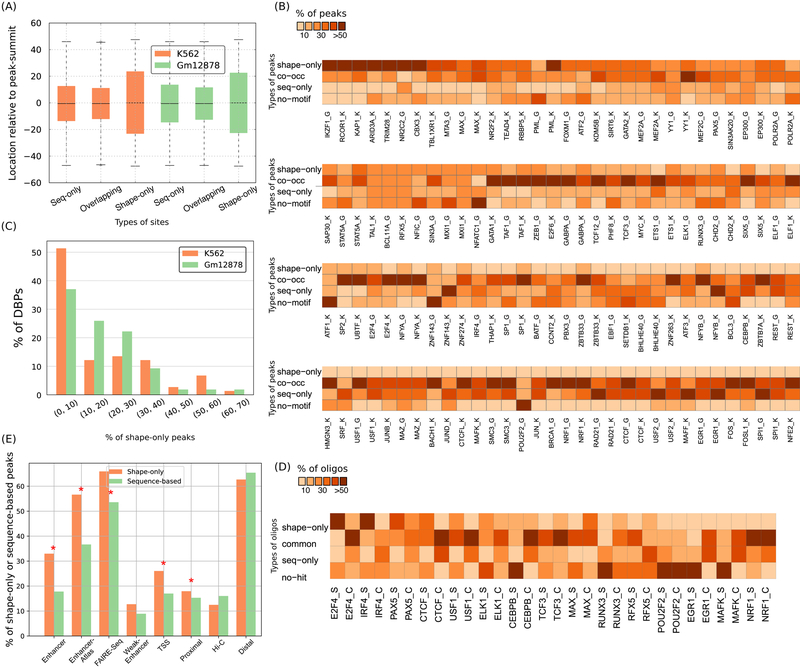
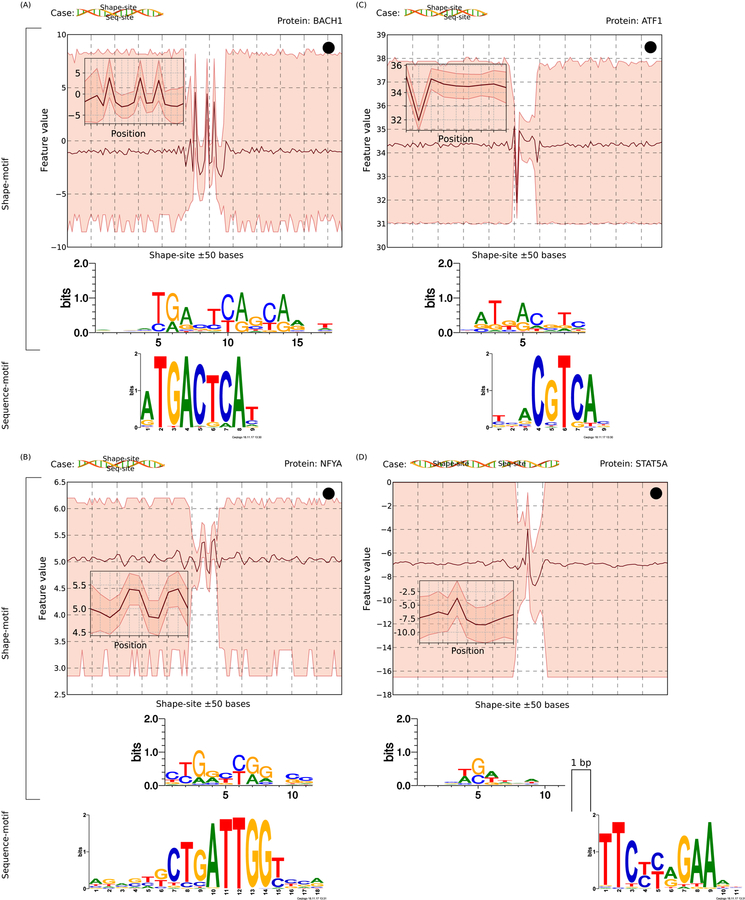
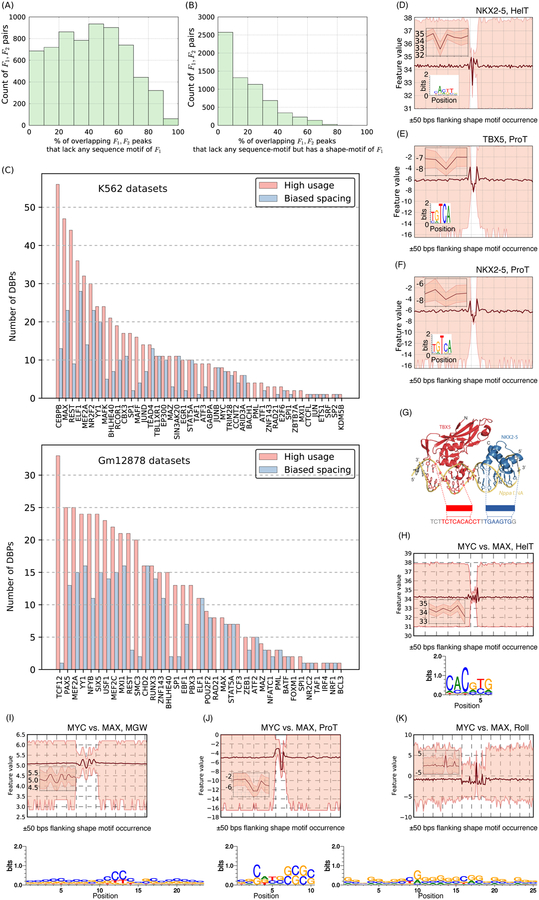
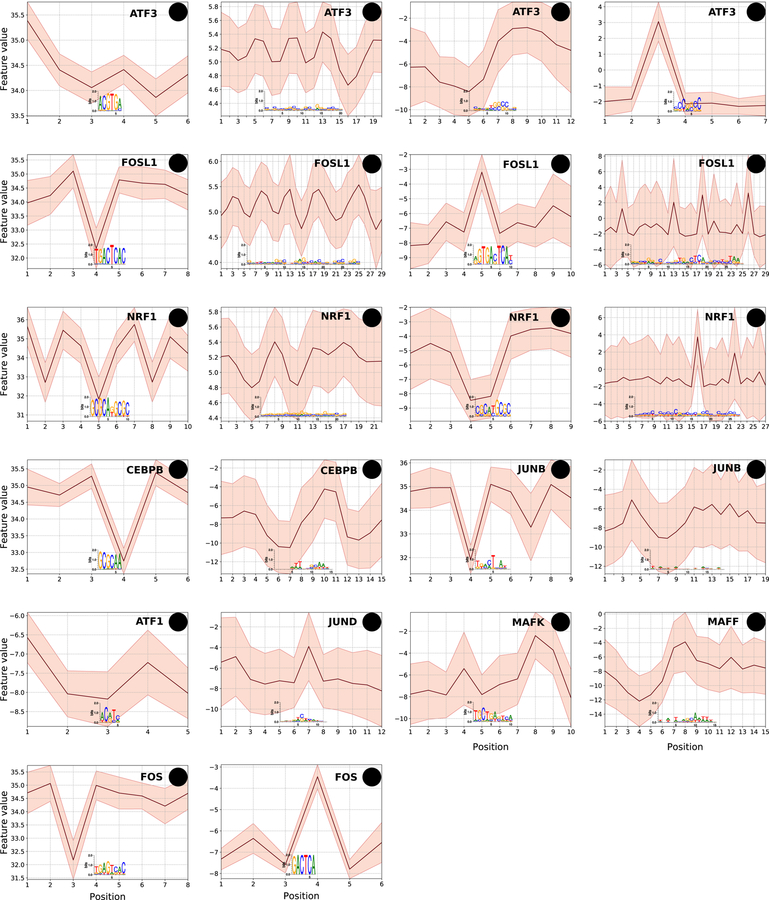
DNA shape adds specificity to sequence motifs but has not been explored systematically outside this context. We hypothesized that DNA-binding proteins (DBPs) preferentially occupy DNA with specific structures ("shape motifs") regardless of whether or not these correspond to high information content sequence motifs. We present ShapeMF, a Gibbs sampling algorithm that identifies de novo shape motifs. Using binding data from hundreds of in vivo and in vitro experiments, we show that most DBPs have shape motifs and can occupy these in the absence of sequence motifs. This "shape-only binding" is common for many DBPs and in regions co-bound by multiple DBPs. When shape and sequence motifs co-occur, they can be overlapping, flanking, or separated by consistent spacing. Finally, DBPs within the same protein family have different shape motifs, explaining their distinct genome-wide occupancy despite having similar sequence motifs. These results suggest that shape motifs not only complement sequence motifs but also facilitate recognition of DNA beyond conventionally defined sequence motifs.
Keywords: ChIP-Seq; DNA binding protein; DNA shape; Gibbs sampling; HT-SELEX; algorithm; de novo shape motif discovery; sequence motif; shape motif; shape specificity; shape-only binding; shape-specific binding; transcription factor.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declarations of Interests
Authors have no financial interest to declare. KSP is a consultant or advisor for Tenaya Therapeutics, uBiome, and Phylagen. BGB is a founder of Tenaya Therapeutics.
Figures
References
-
- Aggarwal AK, Rodgers DW, Drottar M, Ptashne M and Harrison SC (1988). “Recognition of a DNA operator by the repressor of phage 434: a view at high resolution.” Science 242(4880): 899–907. - PubMed
-
- Aishima J and Wolberger C (2003). “Insights into nonspecific binding of homeodomains from a structure of MATalpha2 bound to DNA.” Proteins 51(4): 544–551. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
