

MtABCG20 is an ABA exporter influencing root morphology and seed germination of Medicago truncatula
- PMID: 30661269
- PMCID: PMC6850635
- DOI: 10.1111/tpj.14234
MtABCG20 is an ABA exporter influencing root morphology and seed germination of Medicago truncatula
Abstract
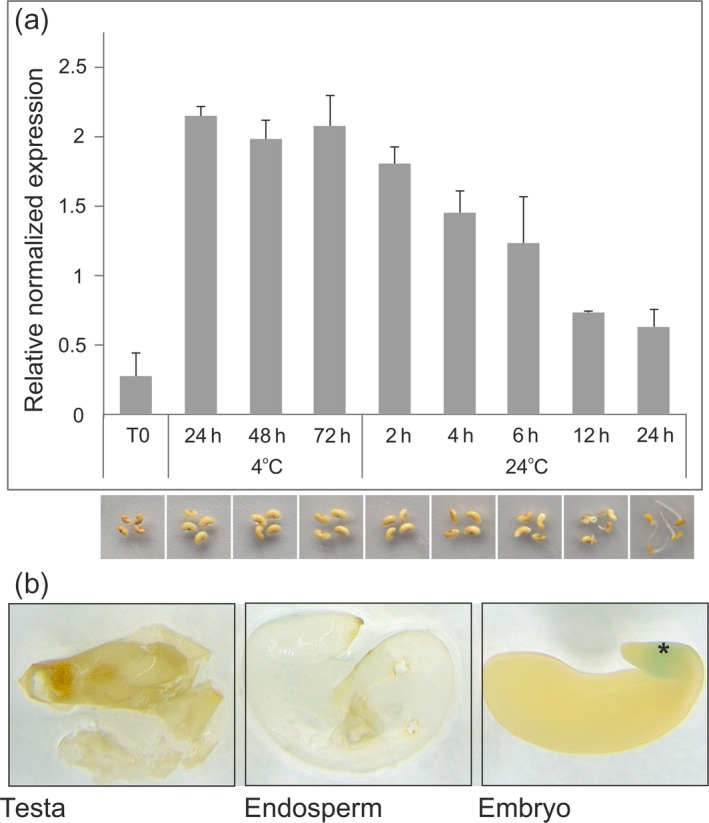
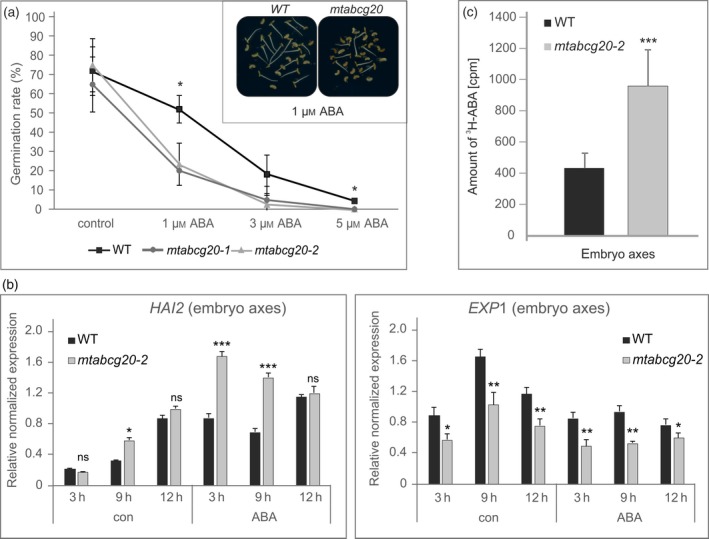
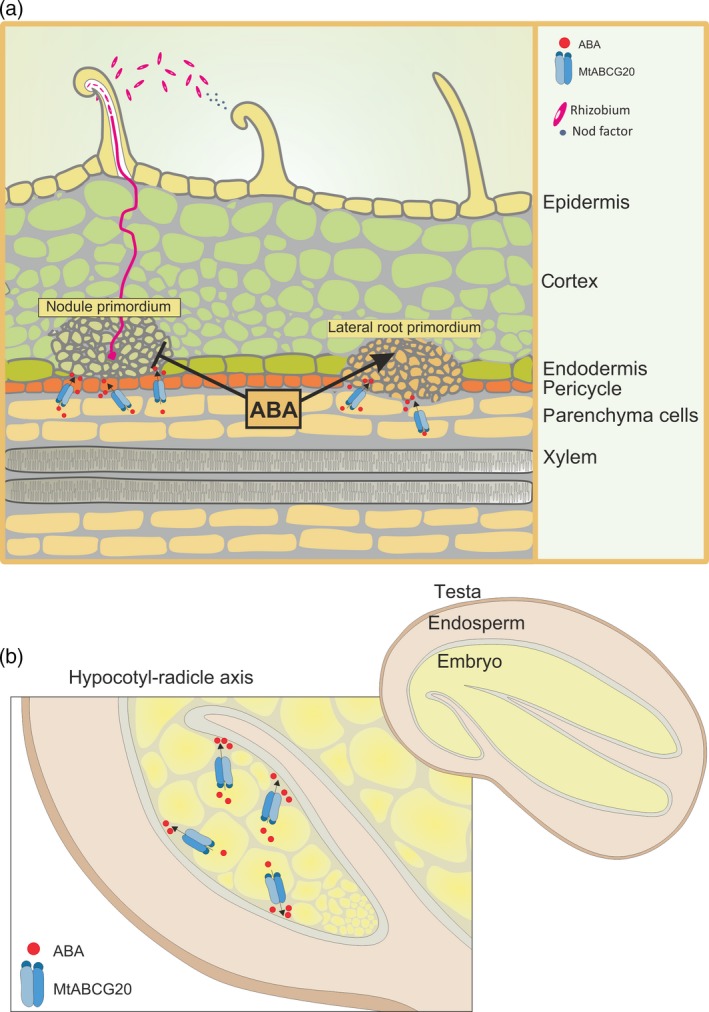
Abscisic acid (ABA) integrates internal and external signals to coordinate plant development, growth and architecture. It plays a central role in stomatal closure, and prevents germination of freshly produced seeds and germination of non-dormant seeds under unfavorable circumstances. Here, we describe a Medicago truncatula ATP-binding cassette (ABC) transporter, MtABCG20, as an ABA exporter present in roots and germinating seeds. In seeds, MtABCG20 was found in the hypocotyl-radicle transition zone of the embryonic axis. Seeds of mtabcg20 plants were more sensitive to ABA upon germination, due to the fact that ABA translocation within mtabcg20 embryos was impaired. Additionally, the mtabcg20 produced fewer lateral roots and formed more nodules compared with wild-type plants in conditions mimicking drought stress. Heterologous expression in Arabidopsis thaliana provided evidence that MtABCG20 is a plasma membrane protein that is likely to form homodimers. Moreover, export of ABA from Nicotiana tabacum BY2 cells expressing MtABCG20 was faster than in the BY2 without MtABCG20. Our results have implications both in legume crop research and determination of the fundamental molecular processes involved in drought response and germination.
Keywords: ABC transporters; abscisic acid; germination; legumes; root organ formation.
© 2019 The Authors. The Plant Journal published by John Wiley & Sons Ltd and Society for Experimental Biology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
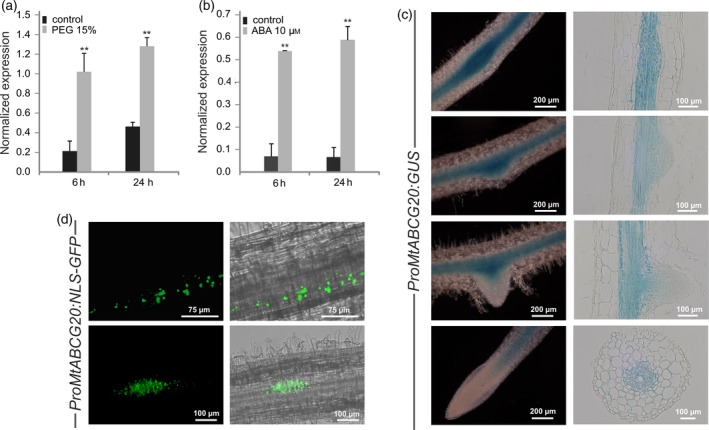
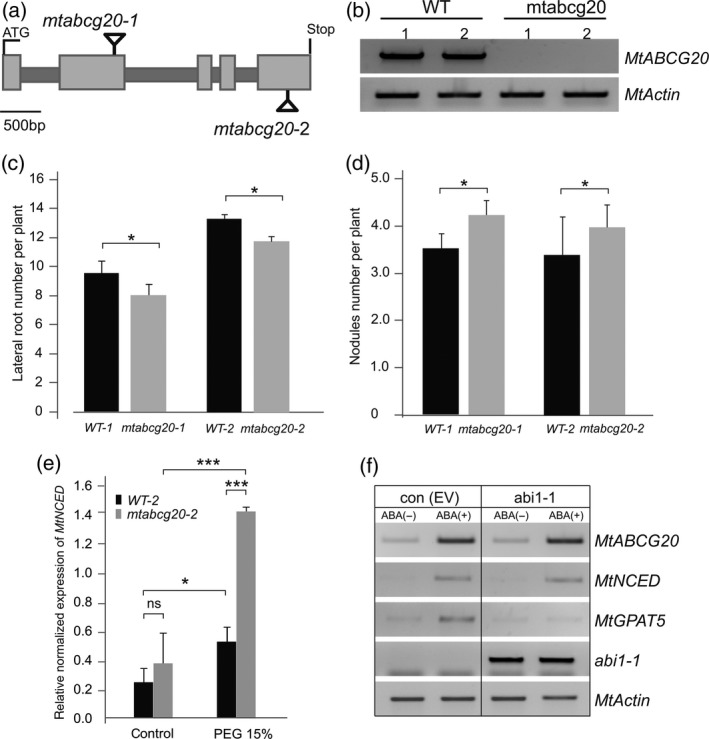
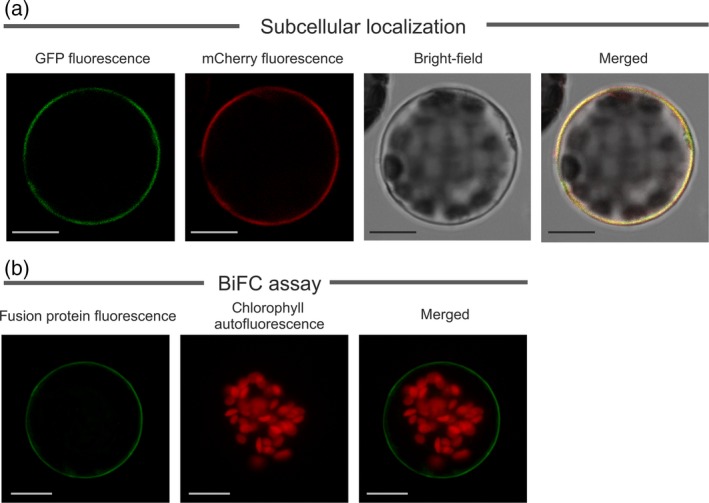
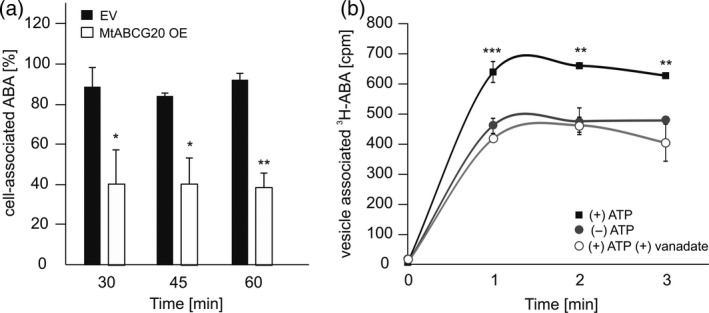
References
-
- Bassel, G.W. , Stamm, P. , Mosca, G. , Barbier de Reuille, P. , Gibbs, D.J. , Winter, R. , Janka, A. , Holdsworth, M.J. and Smith, R.S. (2014) Mechanical constraints imposed by 3D cellular geometry and arrangement modulate growth patterns in the Arabidopsis embryo. Proc. Natl Acad. Sci. USA 111, 8685–8690. - PMC - PubMed
-
- Bauer, H. , Ache, P. , Lautner, S. et al (2013) The stomatal response to reduced relative humidity requires guard cell‐autonomous ABA synthesis. Curr. Biol. 23, 53–57. - PubMed
-
- Bethke, P.C. and Jones, R.L. (2001) Cell death of barley aleurone protoplasts is mediated by reactive oxygen species. Plant J. 25, 19–29. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
