

De Novo SOX4 Variants Cause a Neurodevelopmental Disease Associated with Mild Dysmorphism
- PMID: 30661772
- PMCID: PMC6369454
- DOI: 10.1016/j.ajhg.2018.12.014
De Novo SOX4 Variants Cause a Neurodevelopmental Disease Associated with Mild Dysmorphism
Erratum in
-
De Novo SOX4 Variants Cause a Neurodevelopmental Disease Associated with Mild Dysmorphism.Am J Hum Genet. 2019 Apr 4;104(4):777. doi: 10.1016/j.ajhg.2019.01.014. Am J Hum Genet. 2019. PMID: 30951678 Free PMC article. No abstract available.
Abstract
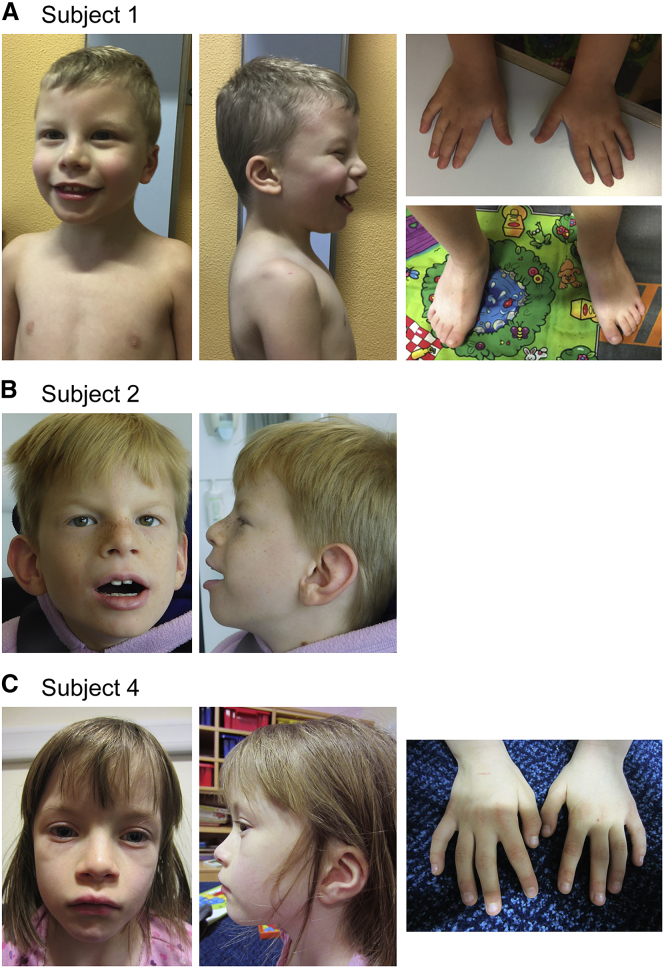
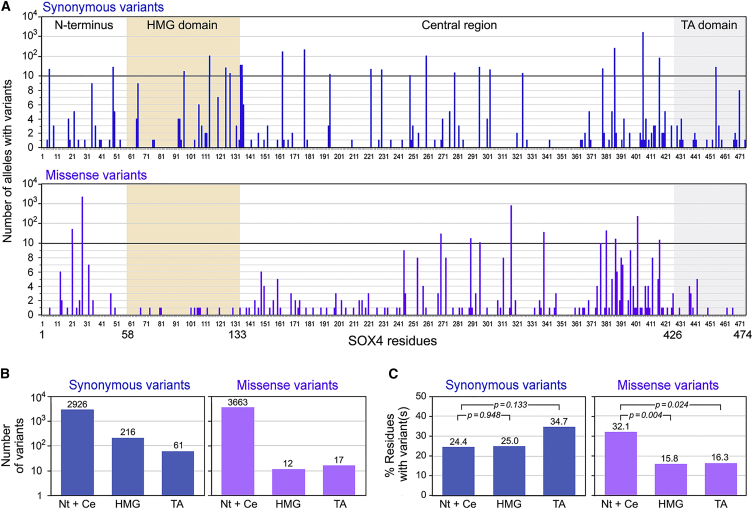
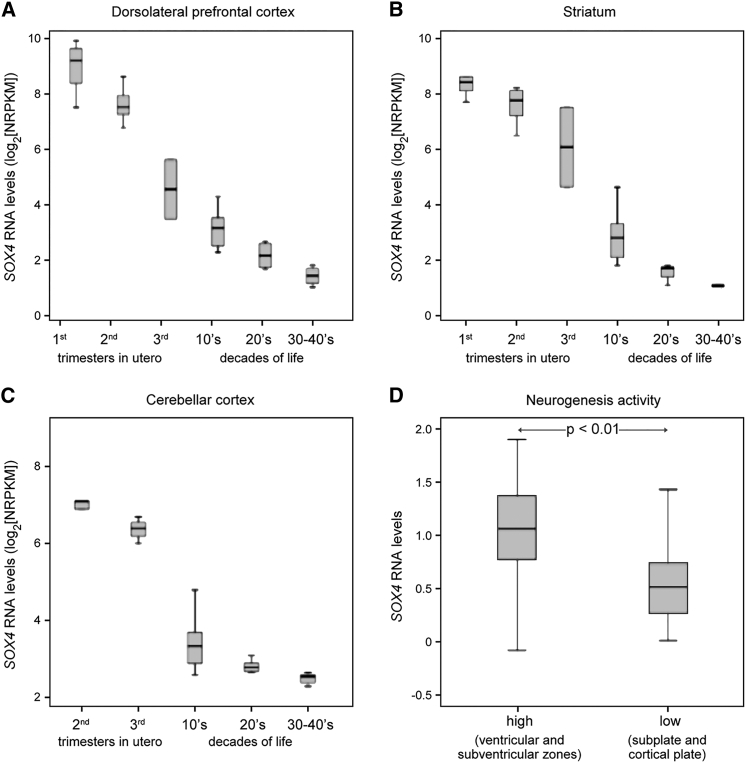
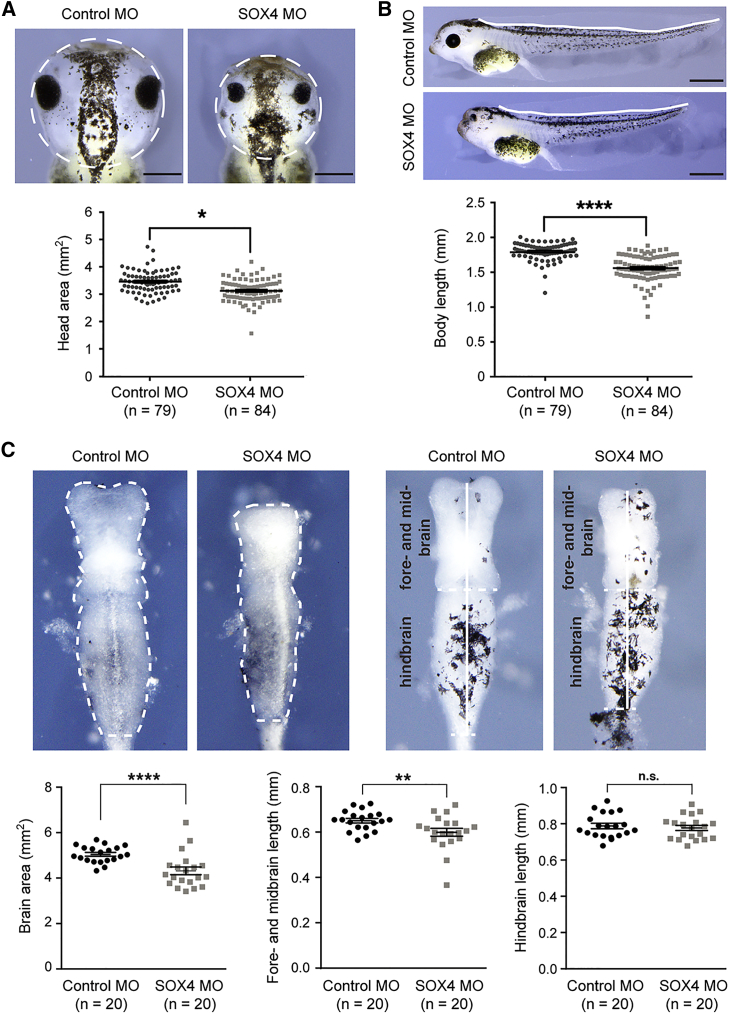
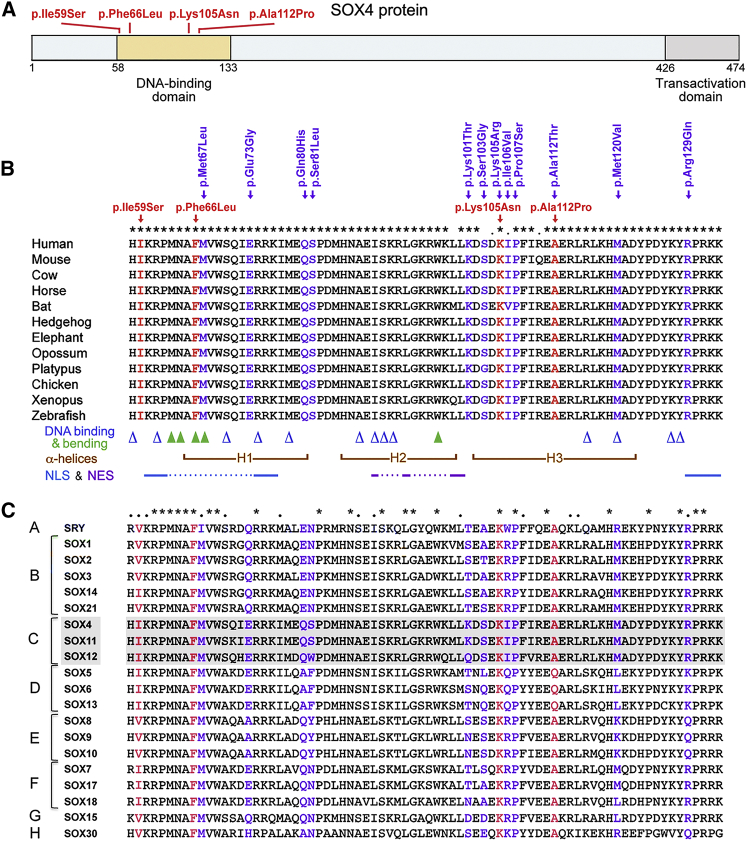
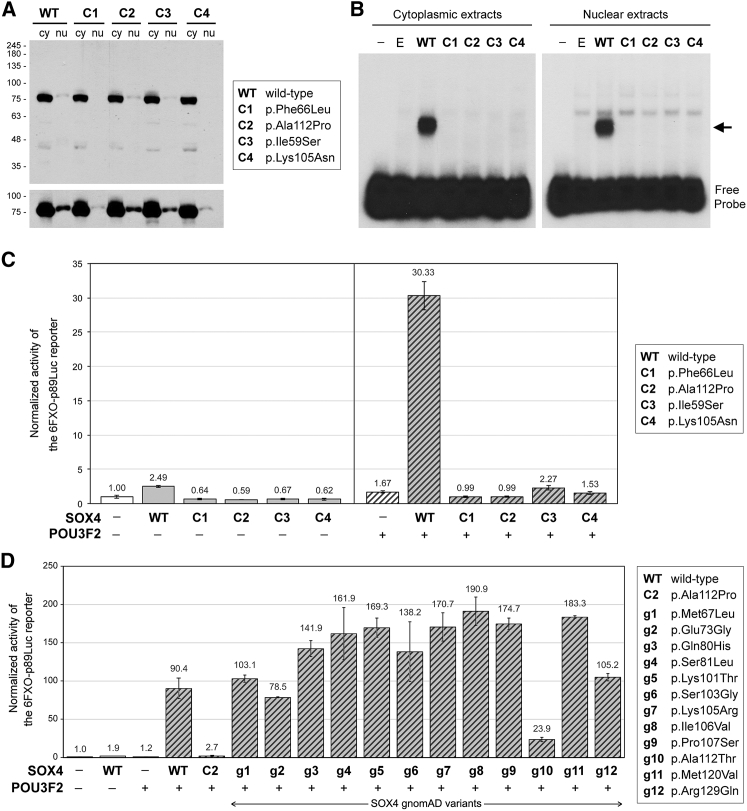
SOX4, together with SOX11 and SOX12, forms group C of SRY-related (SOX) transcription factors. They play key roles, often in redundancy, in multiple developmental pathways, including neurogenesis and skeletogenesis. De novo SOX11 heterozygous mutations have been shown to cause intellectual disability, growth deficiency, and dysmorphic features compatible with mild Coffin-Siris syndrome. Using trio-based exome sequencing, we here identify de novo SOX4 heterozygous missense variants in four children who share developmental delay, intellectual disability, and mild facial and digital morphological abnormalities. SOX4 is highly expressed in areas of active neurogenesis in human fetuses, and sox4 knockdown in Xenopus embryos diminishes brain and whole-body size. The SOX4 variants cluster in the highly conserved, SOX family-specific HMG domain, but each alters a different residue. In silico tools predict that each variant affects a distinct structural feature of this DNA-binding domain, and functional assays demonstrate that these SOX4 proteins carrying these variants are unable to bind DNA in vitro and transactivate SOX reporter genes in cultured cells. These variants are not found in the gnomAD database of individuals with presumably normal development, but 12 other SOX4 HMG-domain missense variants are recorded and all demonstrate partial to full activity in the reporter assay. Taken together, these findings point to specific SOX4 HMG-domain missense variants as the cause of a characteristic human neurodevelopmental disorder associated with mild facial and digital dysmorphism.
Keywords: QA keywords.
Copyright © 2018 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
