

Loss of BCAA Catabolism during Carcinogenesis Enhances mTORC1 Activity and Promotes Tumor Development and Progression
- PMID: 30661928
- PMCID: PMC6506390
- DOI: 10.1016/j.cmet.2018.12.020
Loss of BCAA Catabolism during Carcinogenesis Enhances mTORC1 Activity and Promotes Tumor Development and Progression
Abstract
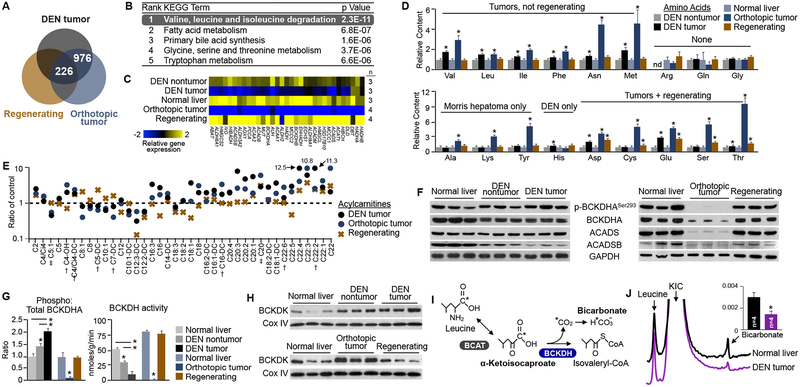
Tumors display profound changes in cellular metabolism, yet how these changes aid the development and growth of tumors is not fully understood. Here we use a multi-omic approach to examine liver carcinogenesis and regeneration, and find that progressive loss of branched-chain amino acid (BCAA) catabolism promotes tumor development and growth. In human hepatocellular carcinomas and animal models of liver cancer, suppression of BCAA catabolic enzyme expression led to BCAA accumulation in tumors, though this was not observed in regenerating liver tissues. The degree of enzyme suppression strongly correlated with tumor aggressiveness, and was an independent predictor of clinical outcome. Moreover, modulating BCAA accumulation regulated cancer cell proliferation in vitro, and tumor burden and overall survival in vivo. Dietary BCAA intake in humans also correlated with cancer mortality risk. In summary, loss of BCAA catabolism in tumors confers functional advantages, which could be exploited by therapeutic interventions in certain cancers.
Keywords: branched-chain amino acids; cancer; cancer metabolism; dietary intake; hepatocellular carcinoma; mTORC1; metabolomics; transcriptomics.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures






References
-
- AICR (2018). Diet, Nutrition, Physical Activity and Cancer: a Global Perspective. Continuous Update Project Expert Report 2018, W.C.R.F.A.I.f.C. Research., ed. (Washington, DC).
-
- An J, Muoio DM, Shiota M, Fujimoto Y, Cline GW, Shulman GI, Koves TR, Stevens R, Millington D, and Newgard CB (2004). Hepatic expression of malonyl-CoA decarboxylase reverses muscle, liver and whole-animal insulin resistance. Nature medicine 10, 268–274. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
