

Organoid-Induced Differentiation of Conventional T Cells from Human Pluripotent Stem Cells
- PMID: 30661959
- PMCID: PMC6687310
- DOI: 10.1016/j.stem.2018.12.011
Organoid-Induced Differentiation of Conventional T Cells from Human Pluripotent Stem Cells
Abstract
The ability to generate T cells from pluripotent stem cells (PSCs) has the potential to transform autologous T cell immunotherapy by facilitating universal, off-the-shelf cell products. However, differentiation of human PSCs into mature, conventional T cells has been challenging with existing methods. We report that a continuous 3D organoid system induced an orderly sequence of commitment and differentiation from PSC-derived embryonic mesoderm through hematopoietic specification and efficient terminal differentiation to naive CD3+CD8αβ+ and CD3+CD4+ conventional T cells with a diverse T cell receptor (TCR) repertoire. Introduction of an MHC class I-restricted TCR in PSCs produced naive, antigen-specific CD8αβ+ T cells that lacked endogenous TCR expression and showed anti-tumor efficacy in vitro and in vivo. Functional assays and RNA sequencing aligned PSC-derived T cells with primary naive CD8+ T cells. The PSC-artificial thymic organoid (ATO) system presented here is an efficient platform for generating functional, mature T cells from human PSCs.
Keywords: 3D organoids; T cell development; conventional T cells; hematopoiesis; human pluripotent stem cells; immunotherapy; in vitro; lymphopoiesis.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
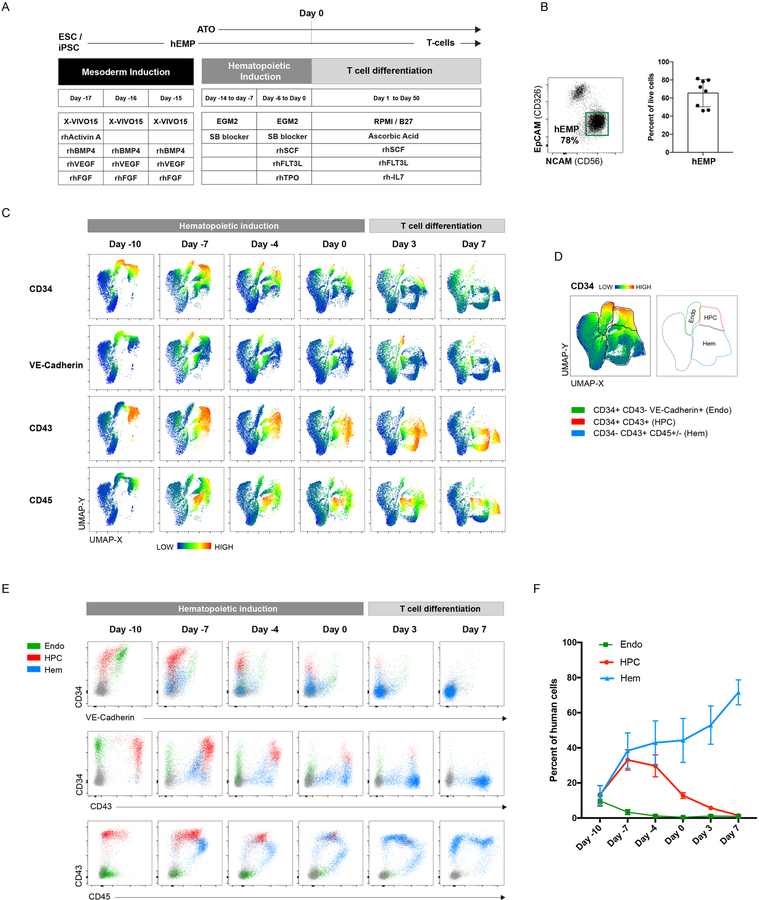
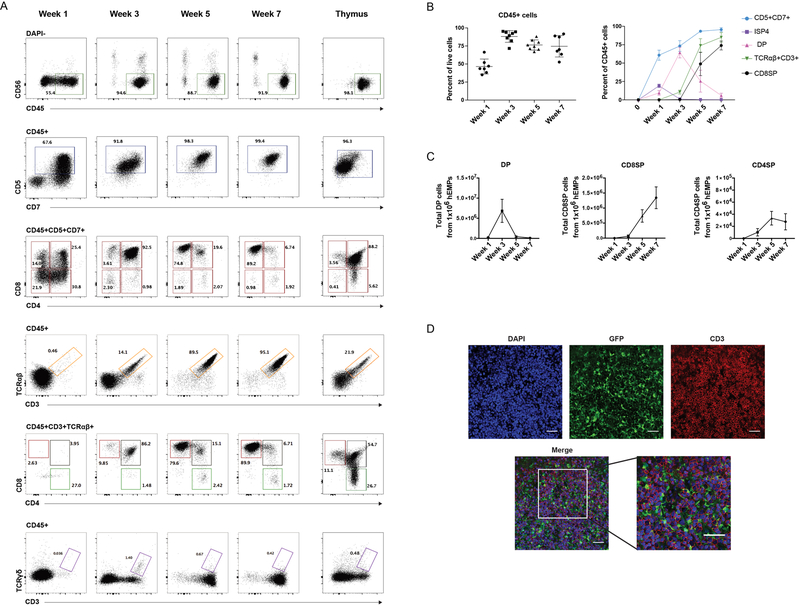
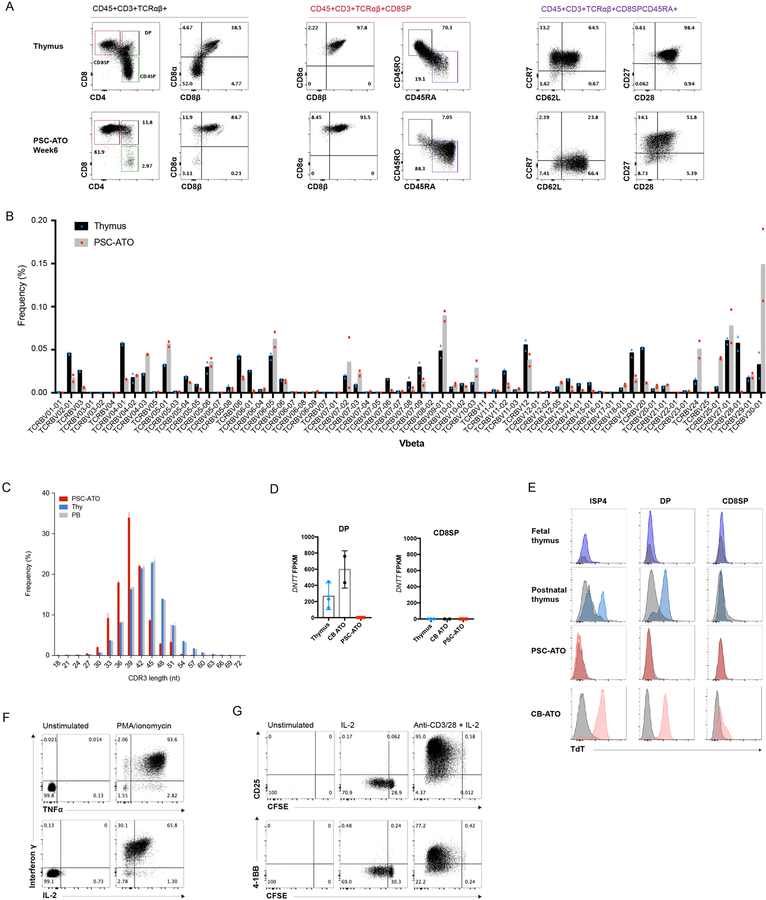
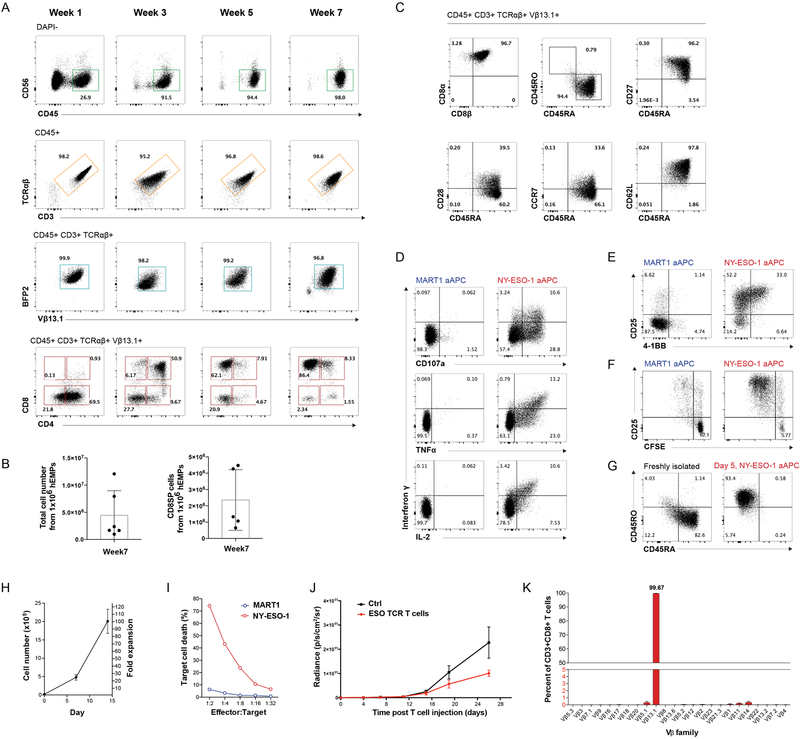
Comment in
-
Close Quarters Can Be a Good Fit for Stem Cells to Become T Cells.Cell Stem Cell. 2019 Mar 7;24(3):345-347. doi: 10.1016/j.stem.2019.02.013. Cell Stem Cell. 2019. PMID: 30849360
References
-
- Crook JM, Peura TT, Kravets L, Bosman AG, Buzzard JJ, Horne R, Hentze H, Dunn NR, Zweigerdt R, Chua F, et al. (2007). The generation of six clinical-grade human embryonic stem cell lines. Cell stem cell 1, 490–494. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
