

The lineage stability and suppressive program of regulatory T cells require protein O-GlcNAcylation
- PMID: 30664665
- PMCID: PMC6341091
- DOI: 10.1038/s41467-019-08300-3
The lineage stability and suppressive program of regulatory T cells require protein O-GlcNAcylation
Abstract
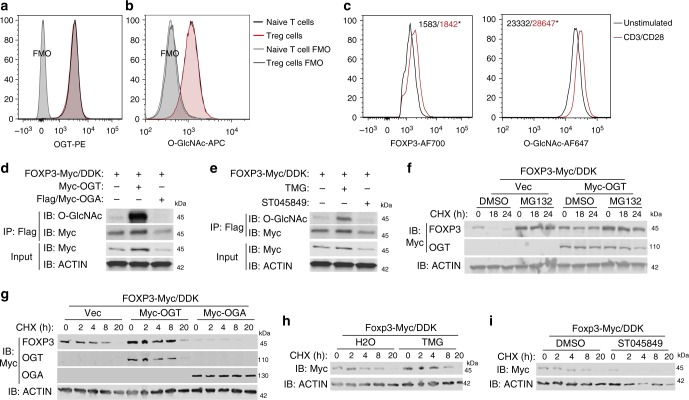
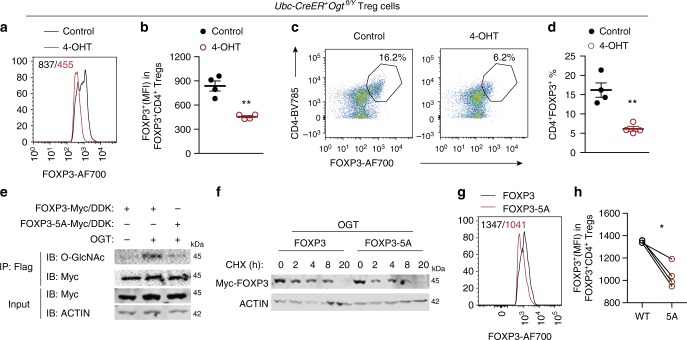
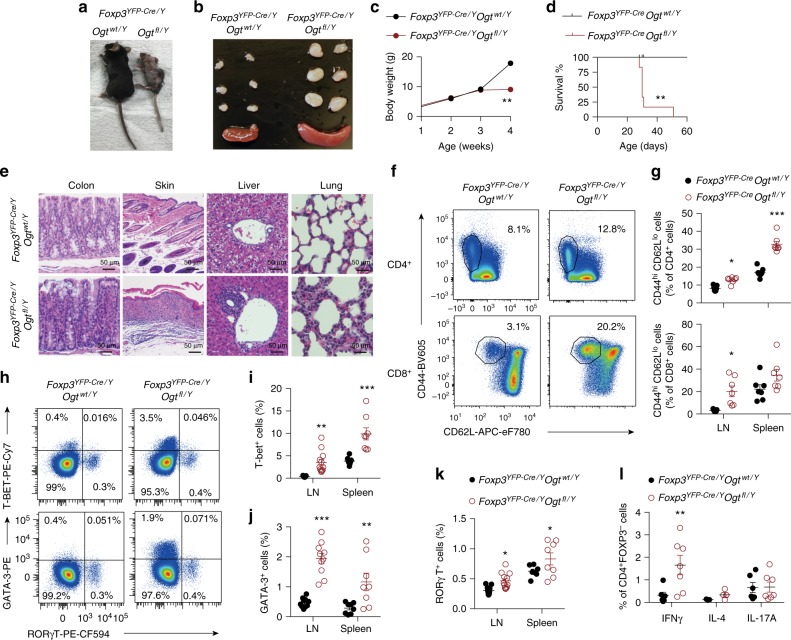
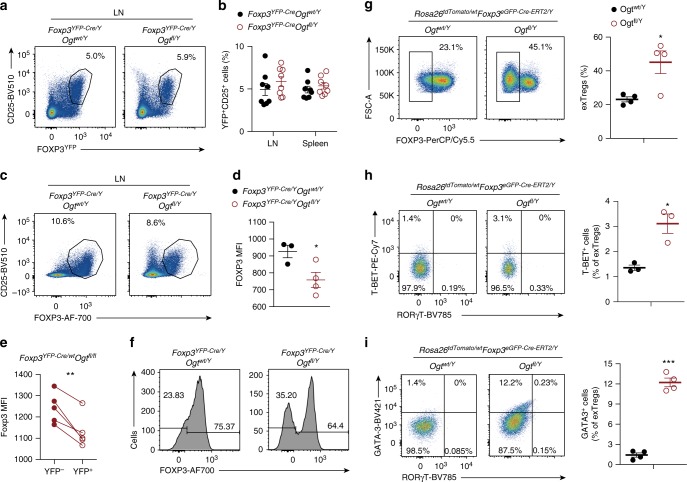
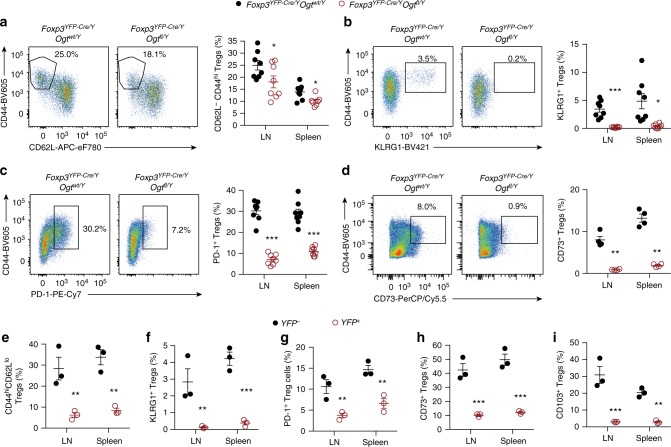
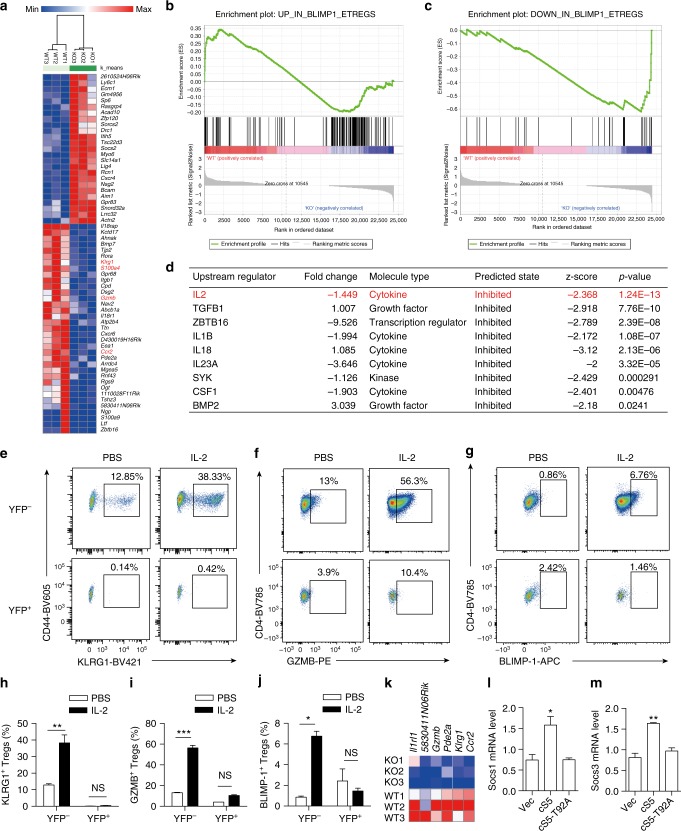
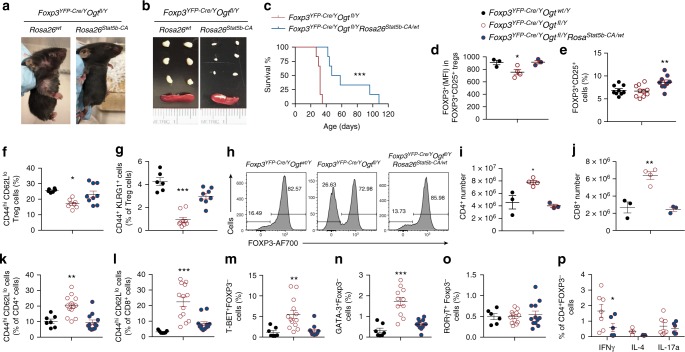
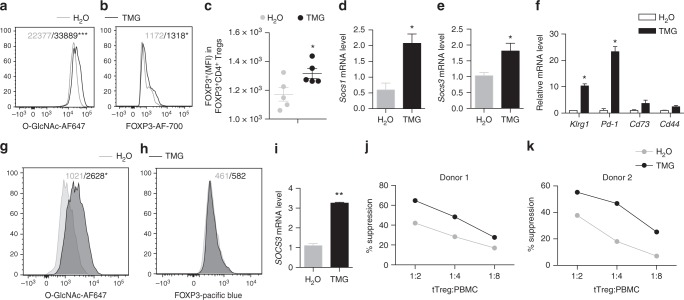
Regulatory T (Treg) cells control self-tolerance, inflammatory responses and tissue homeostasis. In mature Treg cells, continued expression of FOXP3 maintains lineage identity, while T cell receptor (TCR) signaling and interleukin-2 (IL-2)/STAT5 activation support the suppressive effector function of Treg cells, but how these regulators synergize to control Treg cell homeostasis and function remains unclear. Here we show that TCR-activated posttranslational modification by O-linked N-Acetylglucosamine (O-GlcNAc) stabilizes FOXP3 and activates STAT5, thus integrating these critical signaling pathways. O-GlcNAc-deficient Treg cells develop normally but display modestly reduced FOXP3 expression, strongly impaired lineage stability and effector function, and ultimately fatal autoimmunity in mice. Moreover, deficiency in protein O-GlcNAcylation attenuates IL-2/STAT5 signaling, while overexpression of a constitutively active form of STAT5 partially ameliorates Treg cell dysfunction and systemic inflammation in O-GlcNAc deficient mice. Collectively, our data demonstrate that protein O-GlcNAcylation is essential for lineage stability and effector function in Treg cells.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R21 AI140109/AI/NIAID NIH HHS/United States
- P01 CA065493/CA/NCI NIH HHS/United States
- R01 HL118979/HL/NHLBI NIH HHS/United States
- U19 AI051731/AI/NIAID NIH HHS/United States
- R37 AI039560/AI/NIAID NIH HHS/United States
- P01 AI056299/AI/NIAID NIH HHS/United States
- P30 CA077598/CA/NCI NIH HHS/United States
- R01 AI139420/AI/NIAID NIH HHS/United States
- R37 AI034495/AI/NIAID NIH HHS/United States
- R37 HL056067/HL/NHLBI NIH HHS/United States
- R01 HL056067/HL/NHLBI NIH HHS/United States
- S10 OD010731/OD/NIH HHS/United States
- R01 AI124512/AI/NIAID NIH HHS/United States
- P01 CA142106/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
