

Antigenic "Hot- Spots" on the TSH Receptor Hinge Region
- PMID: 30666231
- PMCID: PMC6330735
- DOI: 10.3389/fendo.2018.00765
Antigenic "Hot- Spots" on the TSH Receptor Hinge Region
Abstract
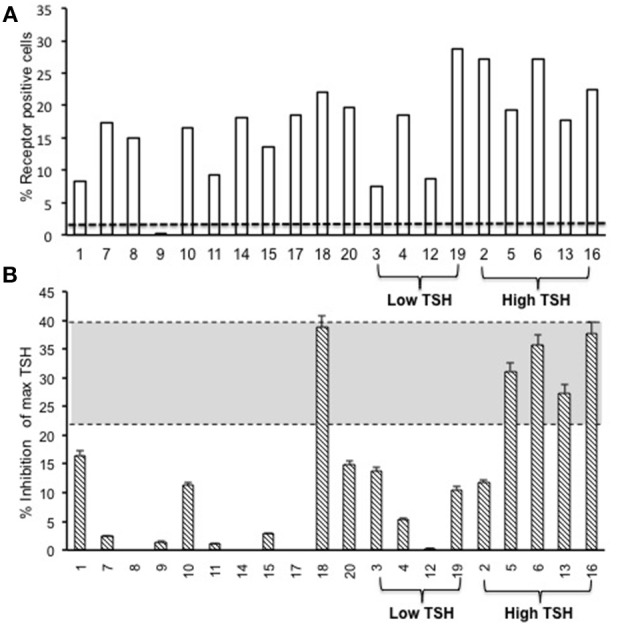
The TSH receptor (TSHR) hinge region was previously considered an inert scaffold connecting the leucine-rich ectodomain to the transmembrane region of the receptor. However, mutation studies have established the hinge region to be an extended hormone-binding site in addition to containing a region which is cleaved thus dividing the receptor into (A) and β (B) subunits. Furthermore, we have shown in-vitro that monoclonal antibodies directed to the cleaved part of the hinge region (often termed "neutral" antibodies) can induce thyroid cell apoptosis in the absence of cyclic AMP signaling. The demonstration of neutral antibodies in patients with Graves' disease suggests their potential involvement in disease pathology thus making the hinge a potentially important antigenic target. Here we examine the evolution of the antibody immune response to the entire TSHR hinge region (aa280-410) after intense immunization with full-length TSHR cDNA in a mouse (BALB/c) model in order to examine the immunogenicity of this critical receptor structure. We found that TSHR hinge region antibodies were detected in 95% of the immunized mice. The antibody responses were largely restricted to residues 352-410 covering three major epitopes and not merely confined to the cleaved portion. These data indicated the presence of novel antigenic "hotspots" within the carboxyl terminus of the hinge region and demonstrate that the hinge region of the TSHR contains an immunogenic pocket that is involved in the highly heterogeneous immune response to the TSHR. The presence of such TSHR antibodies suggests that they may play an active role in the immune repertoire marshaled against the TSHR and may influence the Graves' disease phenotype.
Keywords: Graves' disease (GD); TSH receptor; ectdomain; hinge antibodies; neutral antibodies.
Figures






References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases