

Attenuation of interferon regulatory factor 7 activity in local infectious sites of trachea and lung for preventing the development of acute lung injury caused by influenza A virus
- PMID: 30667045
- PMCID: PMC6459777
- DOI: 10.1111/imm.13045
Attenuation of interferon regulatory factor 7 activity in local infectious sites of trachea and lung for preventing the development of acute lung injury caused by influenza A virus
Abstract
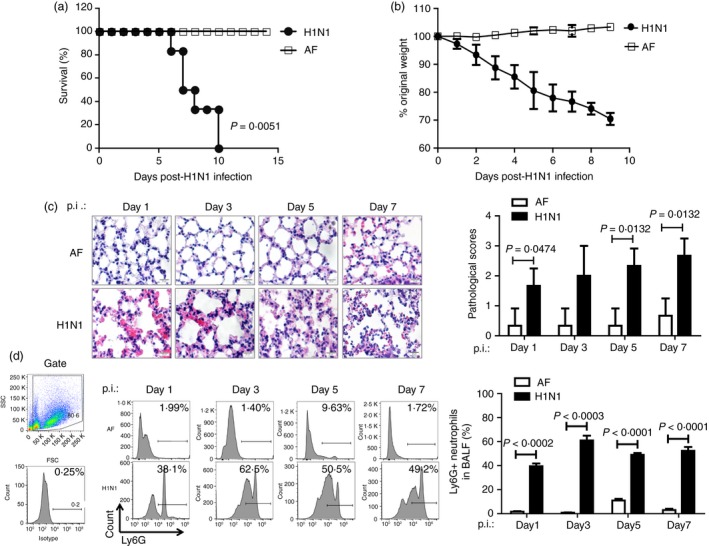
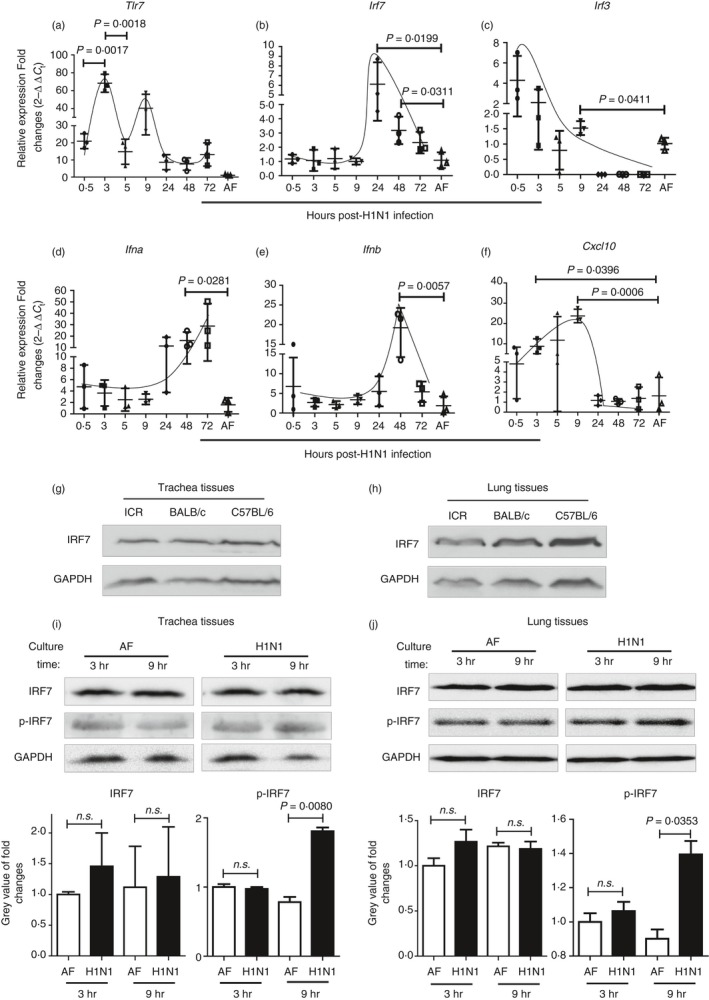
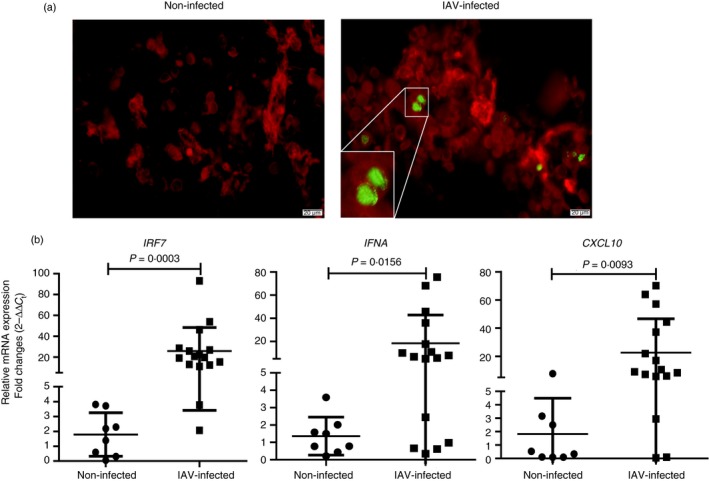
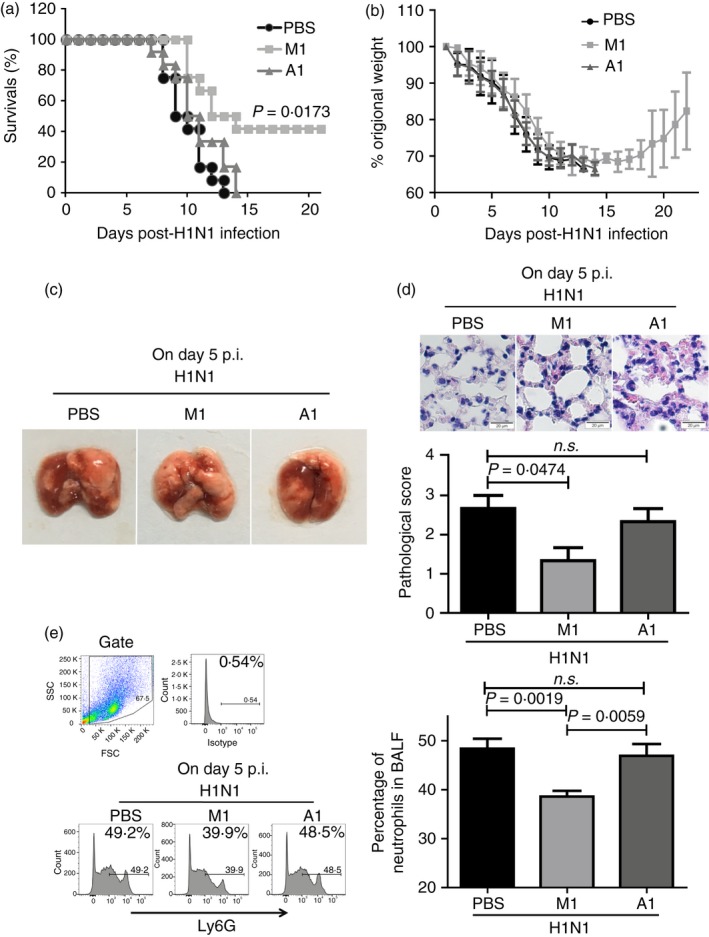
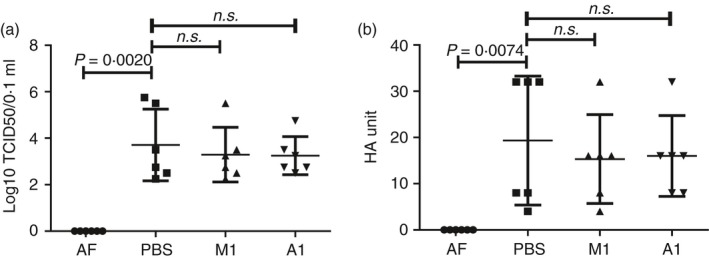
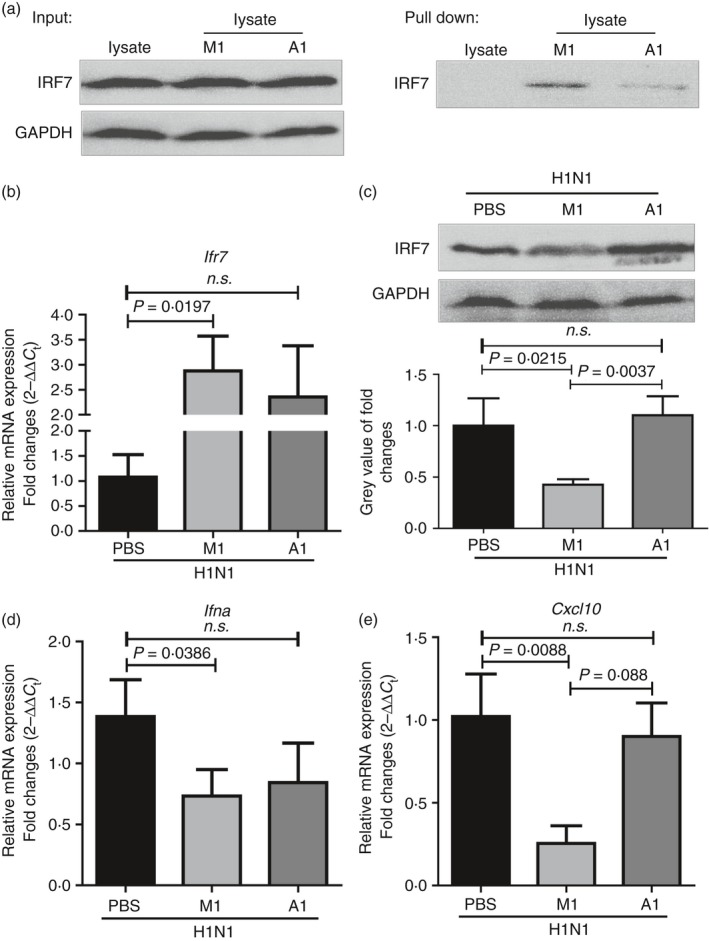
The excessive activation of interferon regulatory factor 7 (IRF7) promotes the development of acute lung injury (ALI) caused by influenza A virus (IAV). However, the deficiency of IRF7 increases the susceptibility to deadly IAV infection in both humans and mice. To test whether the attenuation rather than the abolishment of IRF7 activity in local infectious sites could alleviate IAV-induced ALI, we established IAV-infected mouse model and trachea/lung-tissue culture systems, and designed two IRF7-interfering oligodeoxynucleotides, IRF7-rODN M1 and IRF7-rODN A1, based on the mouse and human consensus sequences of IRF7-binding sites of Ifna/IFNA genes, respectively. In the model mice, we found a close relationship between the IAV-induced ALI and the level/activity of IRF7 in local infectious sites, and also found that the reduced IRF7 level or activity in the lungs of mice treated with IRF7-rODN M1 led to decreased mRNA levels of Ifna genes, reduced neutrophil infiltration in the lungs and prolonged survival of mice. Furthermore, we found that the effects of IRF7-rODN M1 on alleviating IAV-induced ALI could be correlated to the reduced translocation of IRF7, caused by the IRF7-rODN M1, from cytosol to nucleus in IAV-infected cells. These data suggest that the proper attenuation of IRF7 activity in local infectious sites could be a novel approach for treating IAV-induced ALI.
Keywords: acute lung injury; influenza A virus; interferon regulatory factor 7; oligodeoxynucleotide.
© 2019 John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest for the work described here.
Figures
Similar articles
-
IRF7 Is Required for the Second Phase Interferon Induction during Influenza Virus Infection in Human Lung Epithelia.Viruses. 2020 Mar 29;12(4):377. doi: 10.3390/v12040377. Viruses. 2020. PMID: 32235406 Free PMC article.
-
Deletion of Irf3 and Irf7 Genes in Mice Results in Altered Interferon Pathway Activation and Granulocyte-Dominated Inflammatory Responses to Influenza A Infection.J Innate Immun. 2017;9(2):145-161. doi: 10.1159/000450705. Epub 2016 Nov 4. J Innate Immun. 2017. PMID: 27811478 Free PMC article.
-
Basic fibroblast growth factor protects against influenza A virus-induced acute lung injury by recruiting neutrophils.J Mol Cell Biol. 2018 Dec 1;10(6):573-585. doi: 10.1093/jmcb/mjx047. J Mol Cell Biol. 2018. PMID: 29121325
-
Indoleamine 2,3-dioxygenase 1 drives epithelial cells ferroptosis in influenza-induced acute lung injury.Redox Biol. 2025 Apr;81:103572. doi: 10.1016/j.redox.2025.103572. Epub 2025 Feb 26. Redox Biol. 2025. PMID: 40023977 Free PMC article. Review.
-
Journey of monocytes and macrophages upon influenza A virus infection.Curr Opin Virol. 2024 Jun;66:101409. doi: 10.1016/j.coviro.2024.101409. Epub 2024 Apr 2. Curr Opin Virol. 2024. PMID: 38564993 Review.
Cited by
-
The Molecular Mechanism of Multiple Organ Dysfunction and Targeted Intervention of COVID-19 Based on Time-Order Transcriptomic Analysis.Front Immunol. 2021 Aug 24;12:729776. doi: 10.3389/fimmu.2021.729776. eCollection 2021. Front Immunol. 2021. PMID: 34504502 Free PMC article.
-
IRF7 Activates LCN2 Transcription to Enhance LPS-Induced Acute Lung Injury by Inducing Macrophage Ferroptosis and M1 Polarization.Cell Biochem Biophys. 2025 Jun;83(2):2415-2430. doi: 10.1007/s12013-024-01651-9. Epub 2024 Dec 31. Cell Biochem Biophys. 2025. PMID: 39738844
-
The multiple roles of interferon regulatory factor family in health and disease.Signal Transduct Target Ther. 2024 Oct 9;9(1):282. doi: 10.1038/s41392-024-01980-4. Signal Transduct Target Ther. 2024. PMID: 39384770 Free PMC article. Review.
-
Rae1 drives NKG2D binding-dependent tumor development in mice by activating mTOR and STAT3 pathways in tumor cells.Cancer Sci. 2020 Jul;111(7):2234-2247. doi: 10.1111/cas.14434. Epub 2020 May 16. Cancer Sci. 2020. PMID: 32333709 Free PMC article.
-
Molecular markers of type II alveolar epithelial cells in acute lung injury by bioinformatics analysis.Sci Rep. 2023 Oct 18;13(1):17797. doi: 10.1038/s41598-023-45129-9. Sci Rep. 2023. PMID: 37853056 Free PMC article.
References
-
- Damjanovic D, Small CL, Jeyanathan M, McCormick S, Xing Z. Immunopathology in influenza virus infection: uncoupling the friend from foe. Clin Immunol 2012; 144:57–69. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
