

CRISPR/Cas9-Mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance
- PMID: 30669982
- PMCID: PMC6341727
- DOI: 10.1186/s12870-018-1627-4
CRISPR/Cas9-Mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance
Abstract
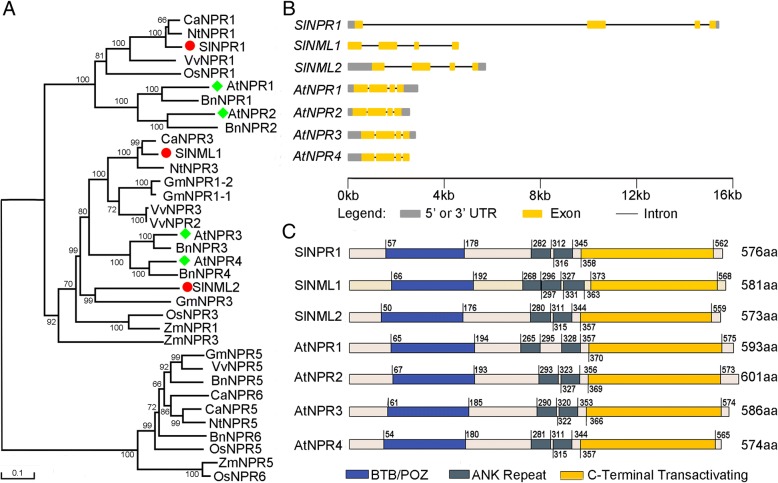
Background: NPR1, nonexpressor of pathogenesis-related gene 1, is a master regulator involved in plant defense response to pathogens, and its regulatory mechanism in the defense pathway has been relatively clear. However, information about the function of NPR1 in plant response to abiotic stress is still limited. Tomato is the fourth most economically crop worldwide and also one of the best-characterized model plants employed in genetic studies. Because of the lack of a stable tomato NPR1 (SlNPR1) mutant, little is known about the function of SlNPR1 in tomato response to biotic and abiotic stresses.
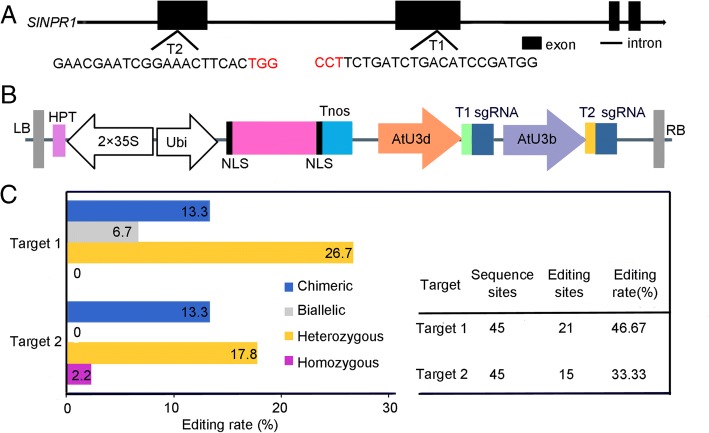
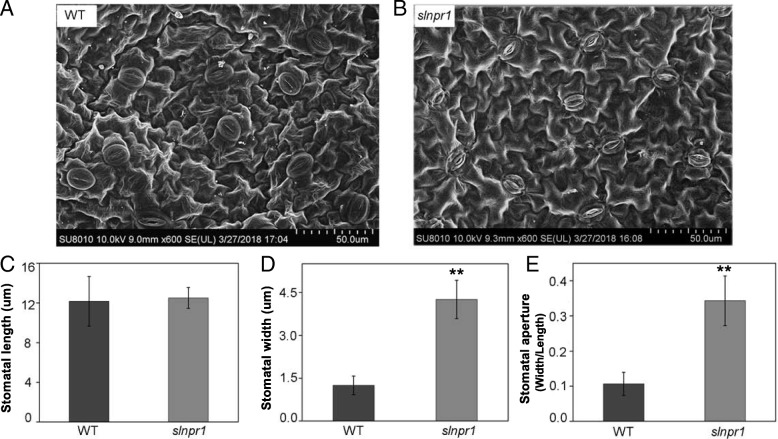
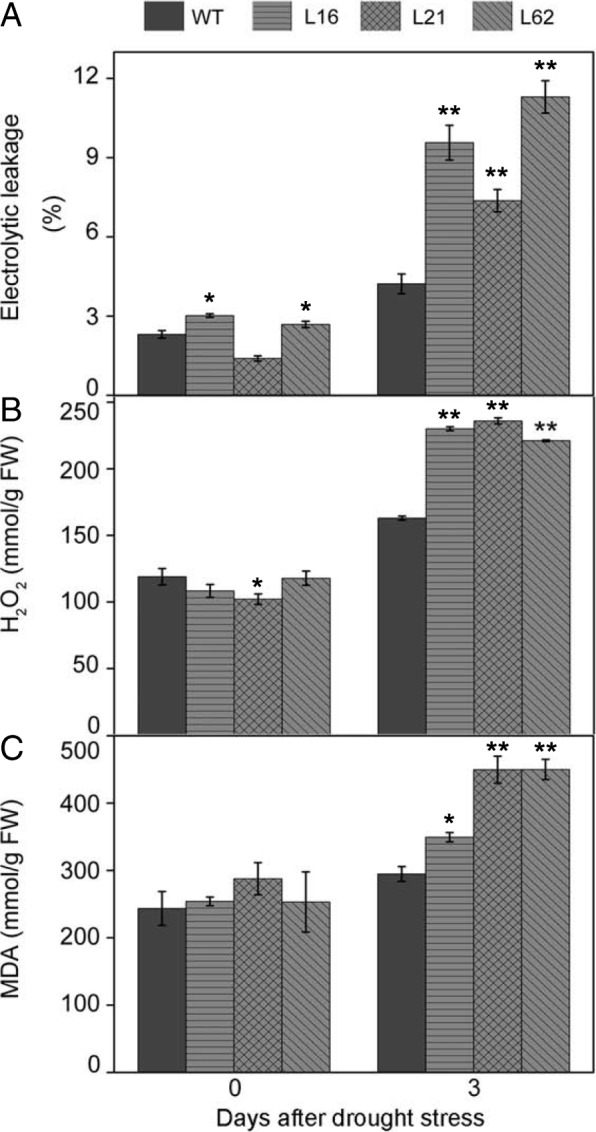
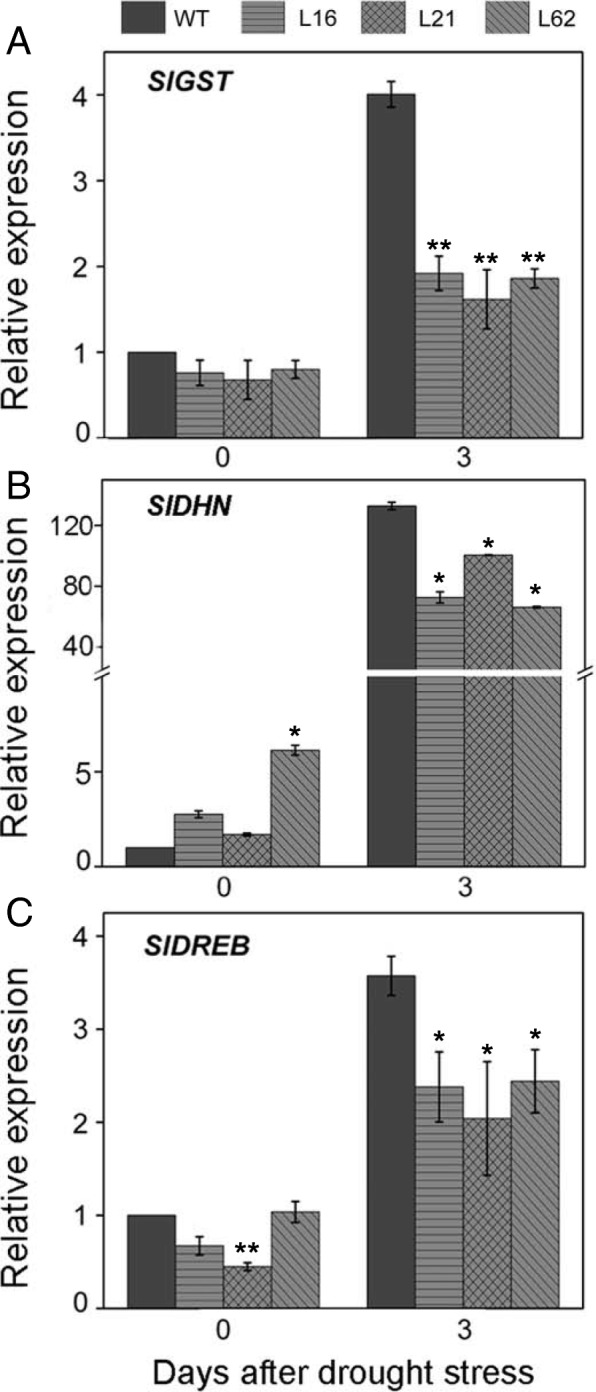
Results: Here we isolated SlNPR1 from tomato 'Ailsa Craig' and generated slnpr1 mutants using the CRISPR/Cas9 system. Analysis of the cis-acting elements indicated that SlNPR1 might be involved in tomato plant response to drought stress. Expression pattern analysis showed that SlNPR1 was expressed in all plant tissues, and it was strongly induced by drought stress. Thus, we investigated the function of SlNPR1 in tomato-plant drought tolerance. Results showed that slnpr1 mutants exhibited reduced drought tolerance with increased stomatal aperture, higher electrolytic leakage, malondialdehyde (MDA) and hydrogen peroxide (H2O2) levels, and lower activity levels of antioxidant enzymes, compared to wild type (WT) plants. The reduced drought tolerance of slnpr1 mutants was further reflected by the down-regulated expression of drought related key genes, including SlGST, SlDHN, and SlDREB.
Conclusions: Collectively, the data suggest that SlNPR1 is involved in regulating tomato plant drought response. These results aid in further understanding the molecular basis underlying SlNPR1 mediation of tomato drought sensitivity.
Keywords: CRISPR/Cas9; Drought; ROS; SlNPR1; Stomatal closure; Tomato plant.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures


References
-
- Mishra KB, Iannacone R, Petrozza A, Mishra A, Armentano N, La Vecchia G, Trtílek M, Cellini F, Nedbal L. Engineered drought tolerance in tomato plants is reflected in chlorophyll fluorescence emission. Plant Sci. 2012;182:79–86. - PubMed
-
- Rai GK, Rai NP, Rathaur S, Kumar S, Singh M. Expression of rd29A::AtDREB1A/CBF3 in tomato alleviates drought-induced oxidative stress by regulating key enzymatic and non-enzymatic antioxidants. Plant Physiol Bioch. 2013;69:90–100. - PubMed
-
- Pantin F, Monnet F, Jannaud D, Costa JM, Renaud J, Muller B, Simonneau T, Genty B. The dual effect of abscisic acid on stomata. New Phytol. 2013;197(1):65–72. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
