

Characterization of l-Carnitine Metabolism in Sinorhizobium meliloti
- PMID: 30670548
- PMCID: PMC6416908
- DOI: 10.1128/JB.00772-18
Characterization of l-Carnitine Metabolism in Sinorhizobium meliloti
Abstract
l-Carnitine is a trimethylammonium compound mostly known for its contribution to fatty acid transport into mitochondria. In bacteria, it is synthesized from γ-butyrobetaine (GBB) and can be used as a carbon source. l-Carnitine can be formed directly by GBB hydroxylation or synthesized via a biosynthetic route analogous to fatty acid degradation. However, this multistep pathway has not been experimentally characterized. In this work, we identified by gene context analysis a cluster of l-carnitine anabolic genes next to those involved in its catabolism and proceeded to the complete in vitro characterization of l-carnitine biosynthesis and degradation in Sinorhizobium meliloti The five enzymes catalyzing the seven steps that convert GBB to glycine betaine are described. Metabolomic analysis confirmed the multistage synthesis of l-carnitine in GBB-grown cells but also revealed that GBB is synthesized by S. meliloti To our knowledge, this is the first report of aerobic GBB synthesis in bacteria. The conservation of l-carnitine metabolism genes in different bacterial taxonomic classes underscores the role of l-carnitine as a ubiquitous nutrient.IMPORTANCE The experimental characterization of novel metabolic pathways is essential for realizing the value of genome sequences and improving our knowledge of the enzymatic capabilities of the bacterial world. However, 30% to 40% of genes of a typical genome remain unannotated or associated with a putative function. We used enzyme kinetics, liquid chromatography-mass spectroscopy (LC-MS)-based metabolomics, and mutant phenotyping for the characterization of the metabolism of l-carnitine in Sinorhizobium meliloti to provide an accurate annotation of the corresponding genes. The occurrence of conserved gene clusters for carnitine metabolism in soil, plant-associated, and marine bacteria underlines the environmental abundance of carnitine and suggests this molecule might make a significant contribution to ecosystem nitrogen and carbon cycling.
Keywords: l-carnitine; LC-MS; bacterial metabolism; enzymology; functional genomics; metabolomics; orbitrap; trimethylammonium compounds.
Copyright © 2019 American Society for Microbiology.
Figures




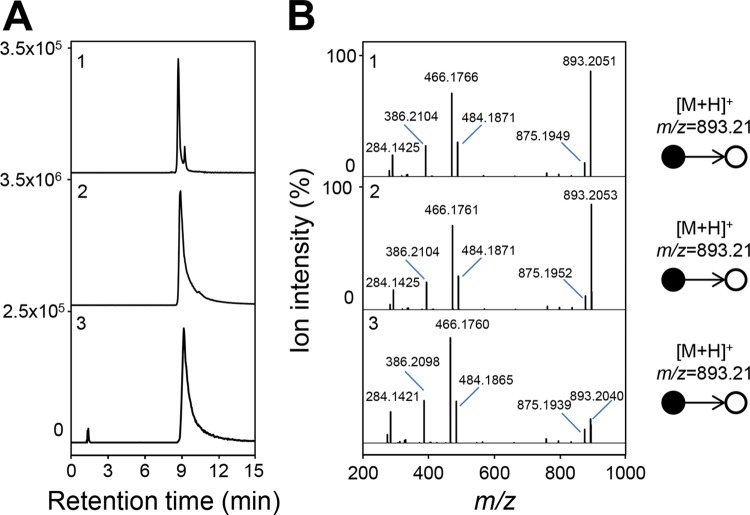
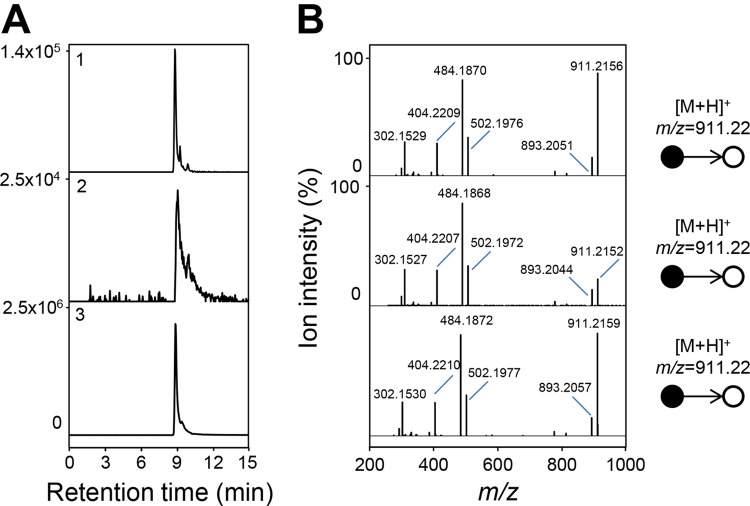
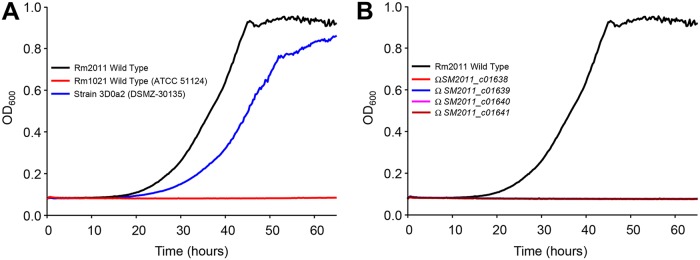
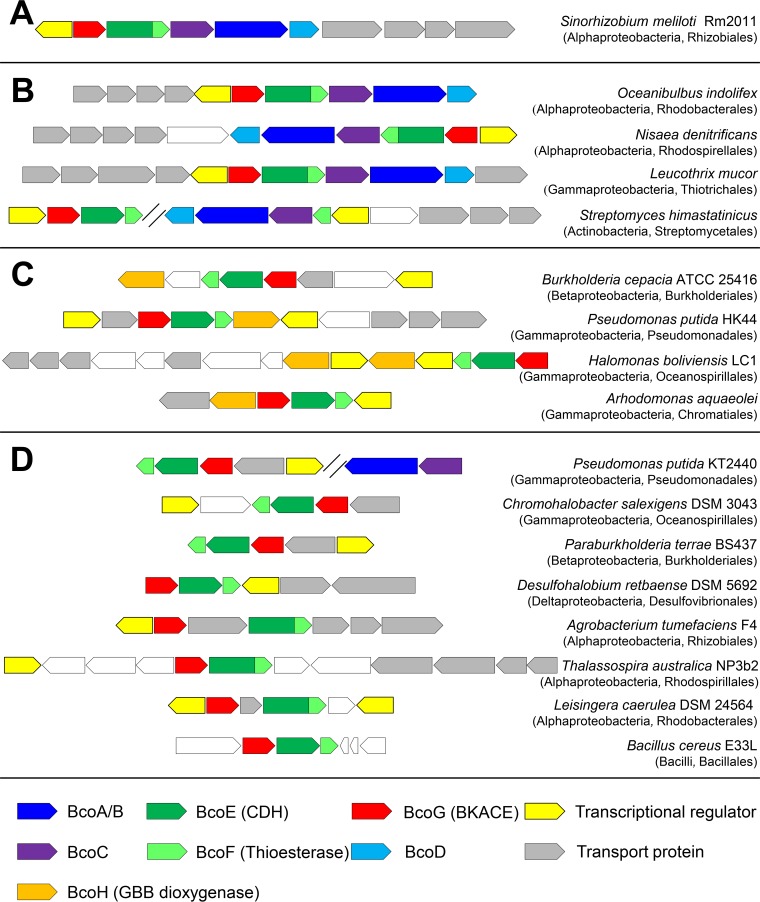
References
-
- Vaz FM, Fouchier SW, Ofman R, Sommer M, Wanders RJ. 2000. Molecular and biochemical characterization of rat gamma-trimethylaminobutyraldehyde dehydrogenase and evidence for the involvement of human aldehyde dehydrogenase 9 in carnitine biosynthesis. J Biol Chem 275:7390–7394. - PubMed
-
- Kaufman RA, Broquist HP. 1977. Biosynthesis of carnitine in Neurospora crassa. J Biol Chem 252:7437–7439. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
