

Diversity of Epigenetic Features of the Inactive X-Chromosome in NK Cells, Dendritic Cells, and Macrophages
- PMID: 30671059
- PMCID: PMC6331414
- DOI: 10.3389/fimmu.2018.03087
Diversity of Epigenetic Features of the Inactive X-Chromosome in NK Cells, Dendritic Cells, and Macrophages
Abstract
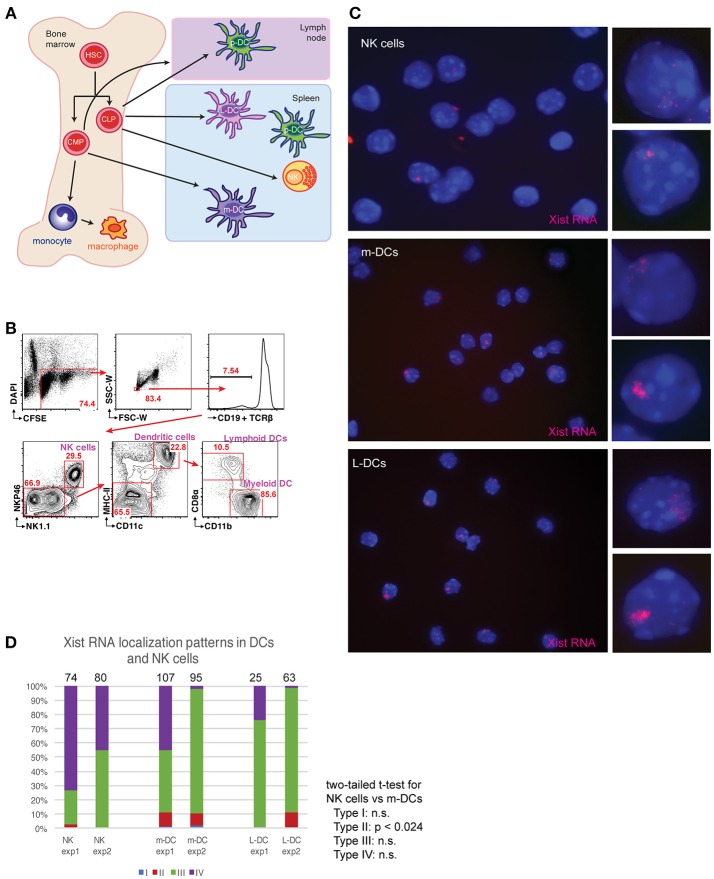
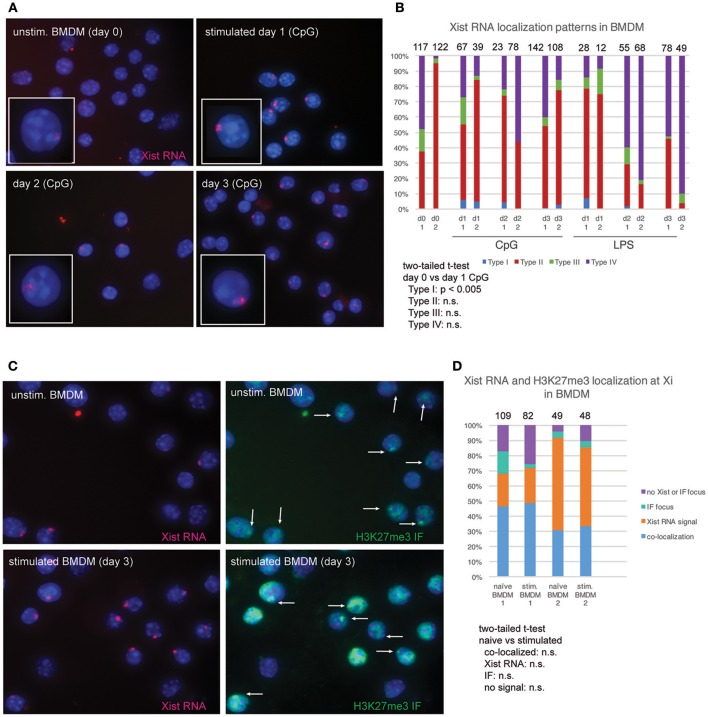
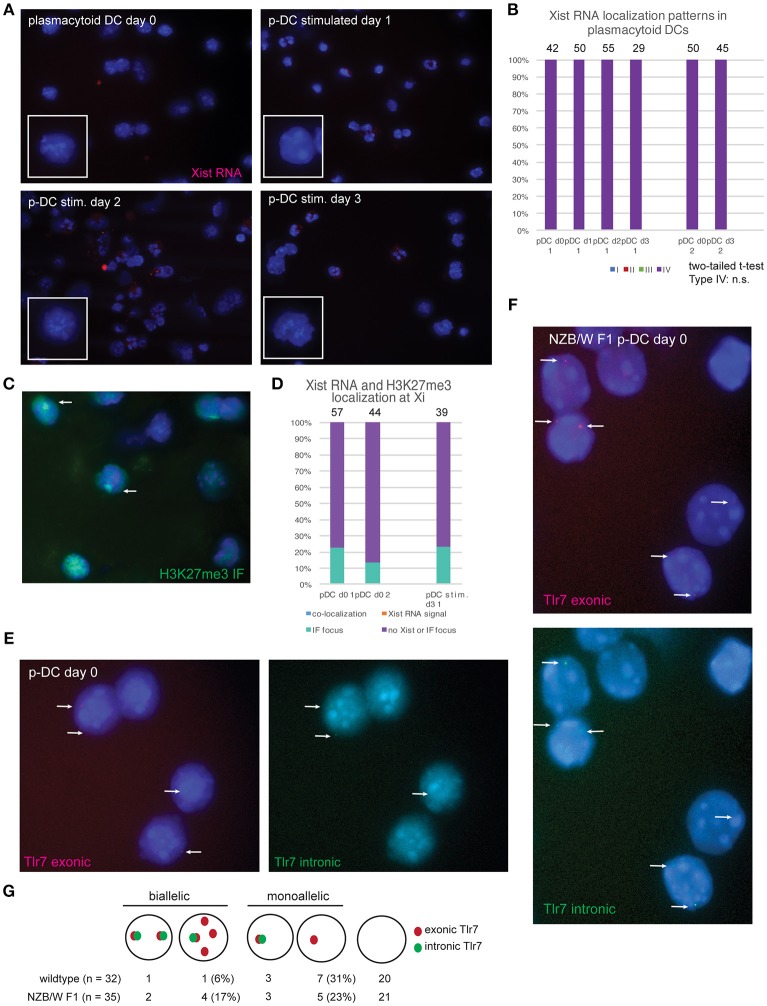
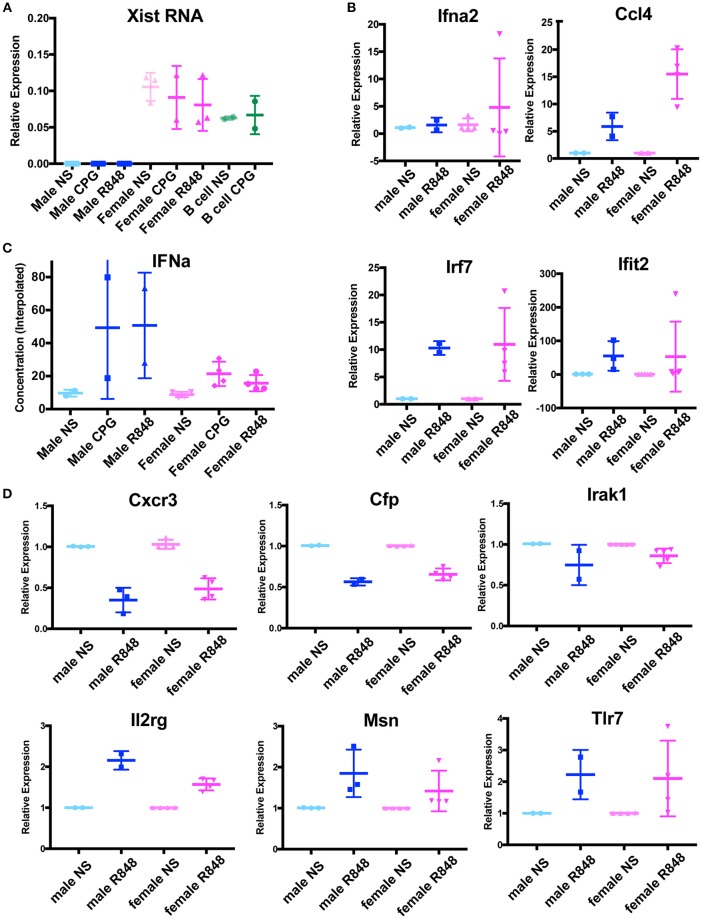
In females, the long non-coding RNA Xist drives X-chromosome Inactivation (XCI) to equalize X-linked gene dosage between sexes. Unlike other somatic cells, dynamic regulation of Xist RNA and heterochromatin marks on the inactive X (Xi) in female lymphocytes results in biallelic expression of some X-linked genes, including Tlr7, Cxcr3, and Cd40l, implicated in sex-biased autoimmune diseases. We now find that while Xist RNA is dispersed across the nucleus in NK cells and dendritic cells (DCs) and partially co-localizes with H3K27me3 in bone marrow-derived macrophages, it is virtually absent in plasmacytoid DCs (p-DCs). Moreover, H3K27me3 foci are present in only 10-20% of cells and we observed biallelic expression of Tlr7 in p-DCs from wildtype mice and NZB/W F1 mice. Unlike in humans, mouse p-DCs do not exhibit sex differences with interferon alpha production, and interferon signature gene expression in p-DCs is similar between males and females. Despite the absence of Xist RNA from the Xi, female p-DCs maintain dosage compensation of six immunity-related X-linked genes. Thus, immune cells use diverse mechanisms to maintain XCI which could contribute to sex-linked autoimmune diseases.
Keywords: NK cells; X-chromosome inactivation; Xist RNA; interferon alpha; long non-coding RNA; macrophages; plasmacytoid dendritic cells; sex differences.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources