

An analog of glibenclamide selectively enhances autophagic degradation of misfolded α1-antitrypsin Z
- PMID: 30673724
- PMCID: PMC6343872
- DOI: 10.1371/journal.pone.0209748
An analog of glibenclamide selectively enhances autophagic degradation of misfolded α1-antitrypsin Z
Abstract
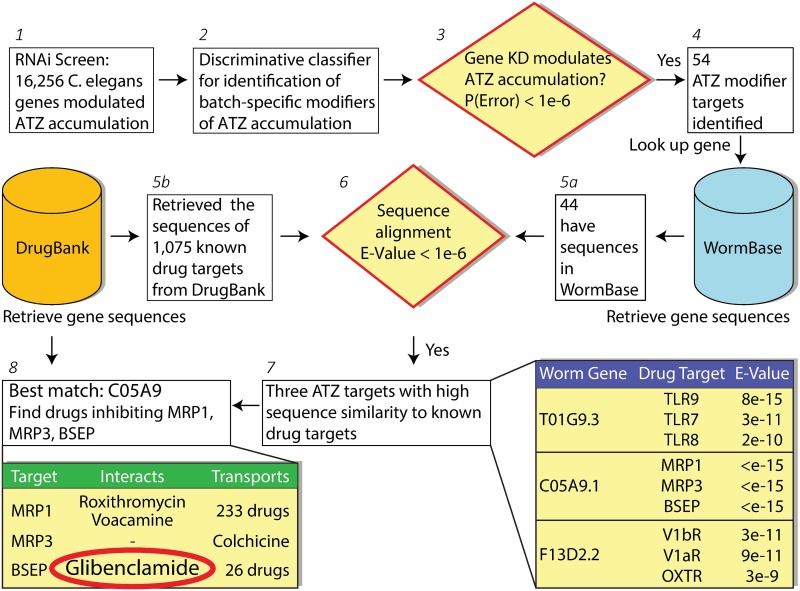
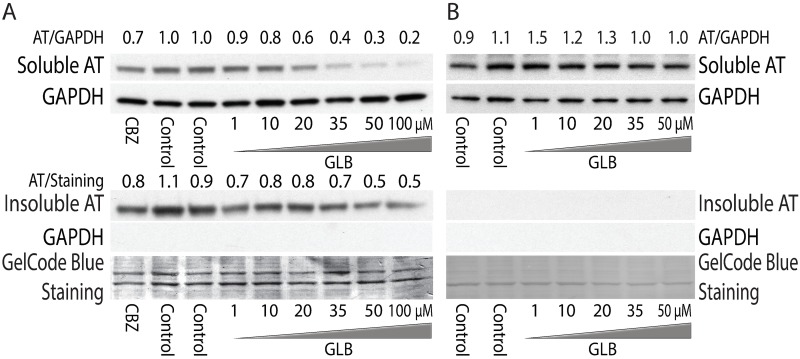
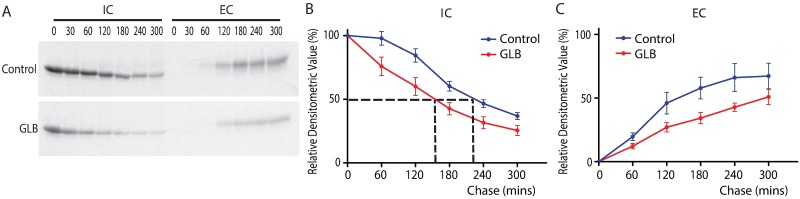
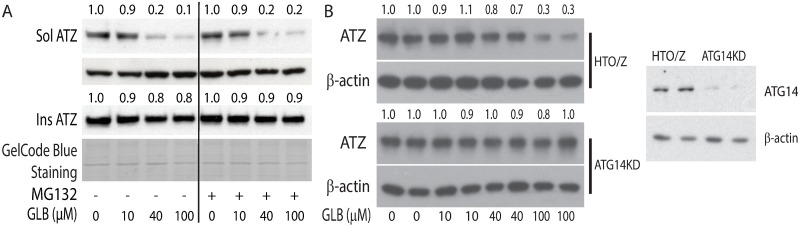
The classical form of α1-antitrypsin deficiency (ATD) is characterized by intracellular accumulation of the misfolded variant α1-antitrypsin Z (ATZ) and severe liver disease in some of the affected individuals. In this study, we investigated the possibility of discovering novel therapeutic agents that would reduce ATZ accumulation by interrogating a C. elegans model of ATD with high-content genome-wide RNAi screening and computational systems pharmacology strategies. The RNAi screening was utilized to identify genes that modify the intracellular accumulation of ATZ and a novel computational pipeline was developed to make high confidence predictions on repurposable drugs. This approach identified glibenclamide (GLB), a sulfonylurea drug that has been used broadly in clinical medicine as an oral hypoglycemic agent. Here we show that GLB promotes autophagic degradation of misfolded ATZ in mammalian cell line models of ATD. Furthermore, an analog of GLB reduces hepatic ATZ accumulation and hepatic fibrosis in a mouse model in vivo without affecting blood glucose or insulin levels. These results provide support for a drug discovery strategy using simple organisms as human disease models combined with genetic and computational screening methods. They also show that GLB and/or at least one of its analogs can be immediately tested to arrest the progression of human ATD liver disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
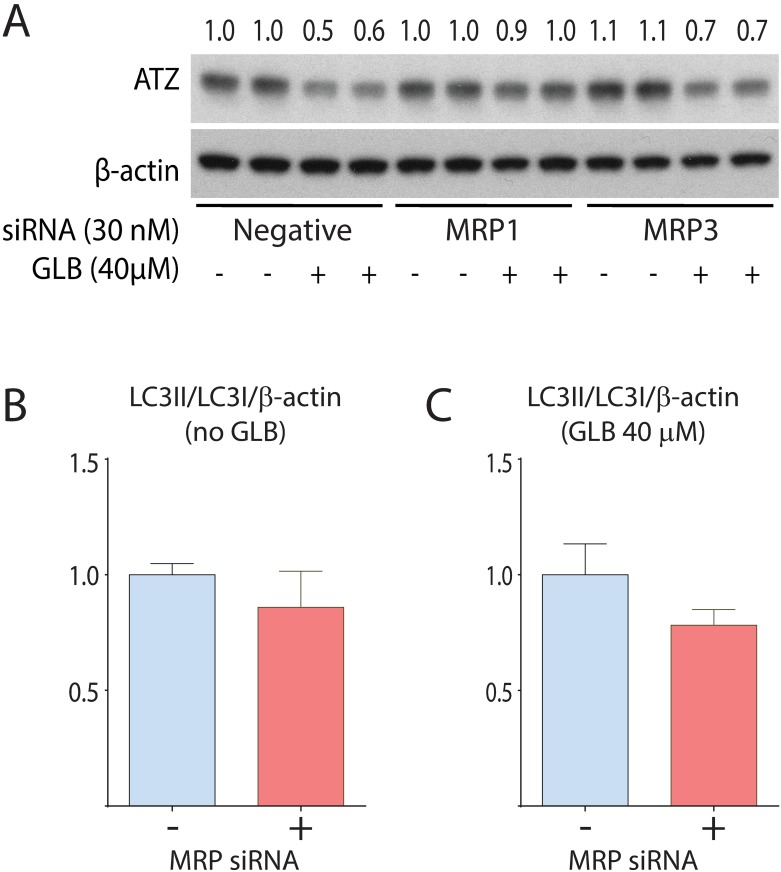
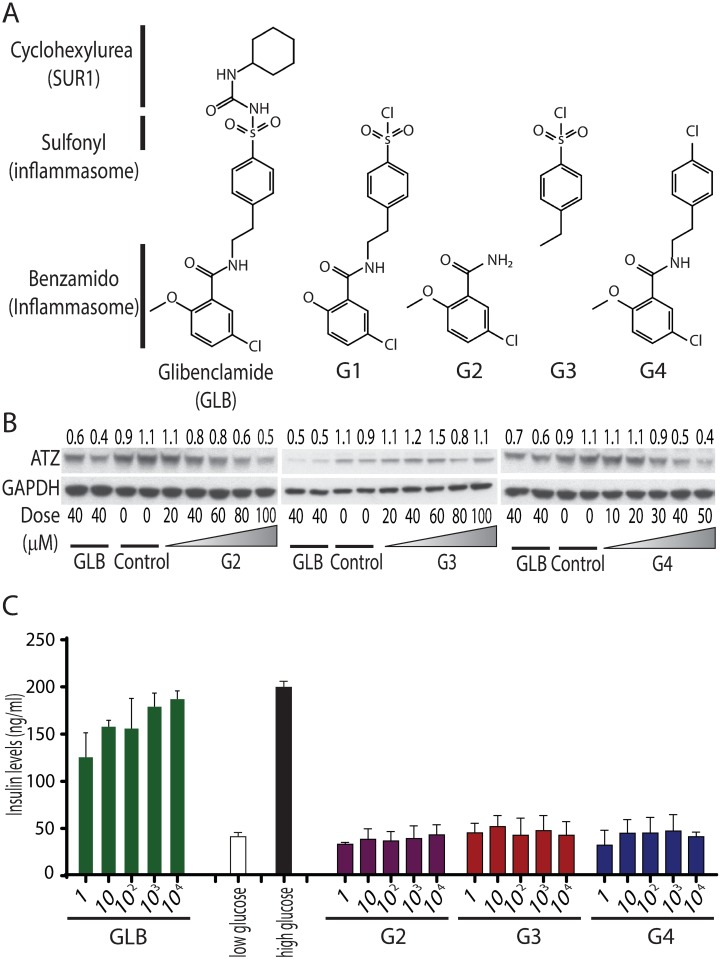
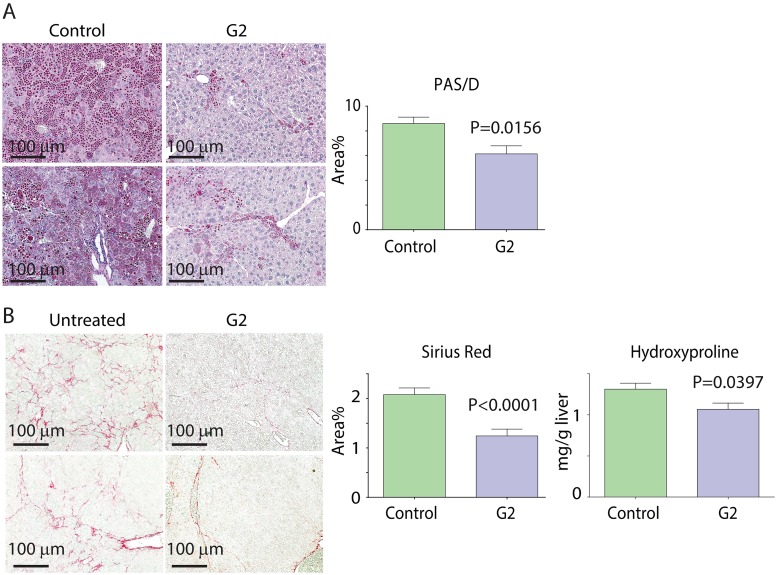
References
-
- Tafaleng EN, Chakraborty S, Han B, Hale P, Wu W, Soto-Gutierrez A, Feghali-Bostwick CA,Wilson AA,Kotton DN, Nagaya M, Strom SC, Roy-Chowdhury J, Stolz DB, Perlmutter DH, Fox IJ (2015). Induced pluripotent stem cells model personalized variations in liver disease resulting from α1-antitrypsin deficiency. Hepatology 62: 147–157. 10.1002/hep.27753 Epub 2015 Apr 13. - DOI - PMC - PubMed
-
- O'Reilly LP, Long OS, Cobanoglu MC, Benson JA, Luke CJ, Miedel MT, Hale P, Perlmutter DH, Bahar I, Silverman GA, Pak SC (2014). A genome-wide RNAi screen identifies potential drug targets in a C. elegans model of alpha1-antitrypsin deficiency. Hum Mol Genet 23:5123–5132. 10.1093/hmg/ddu236 Epub 2014 May 16. - DOI - PMC - PubMed
-
- Li J, Pak SC, O'Reilly LP, Benson JA, Wang Y, Hidvegi T, Hale P, Dippold C, Ewing M, Silverman GA, Perlmutter DH (2014). Fluphenazine reduces proteotoxicity in C. elegans and mammalian models of alpha-1-antitrypsin deficiency. PLoS One 9(1):e87260 10.1371/journal.pone.0087260 eCollection 2014. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
