

An ancient germ cell-specific RNA-binding protein protects the germline from cryptic splice site poisoning
- PMID: 30674417
- PMCID: PMC6345566
- DOI: 10.7554/eLife.39304
An ancient germ cell-specific RNA-binding protein protects the germline from cryptic splice site poisoning
Abstract
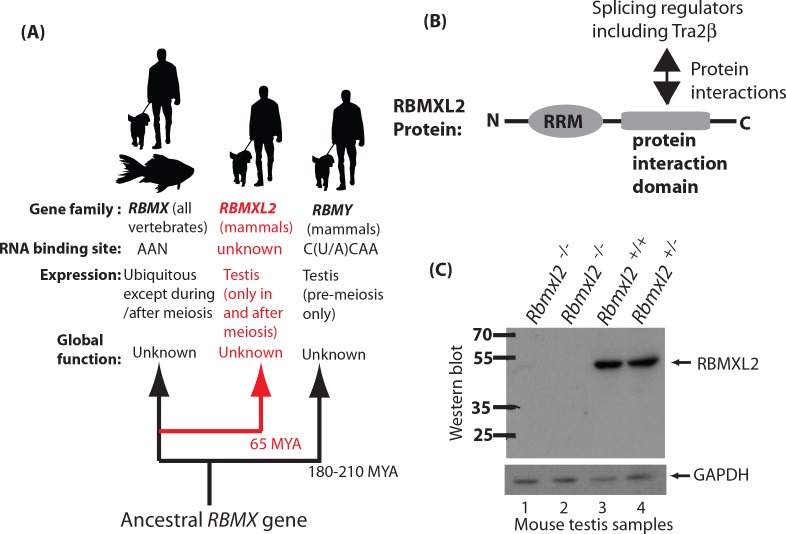
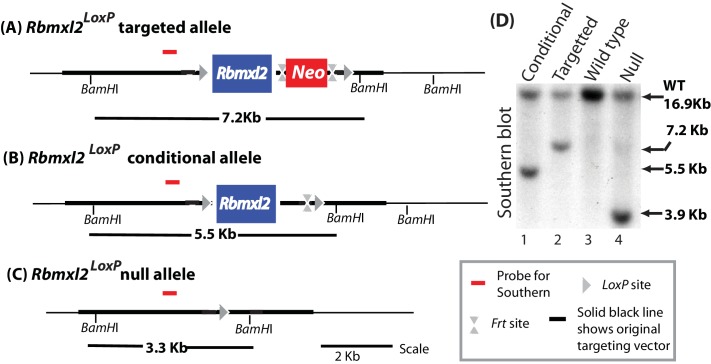
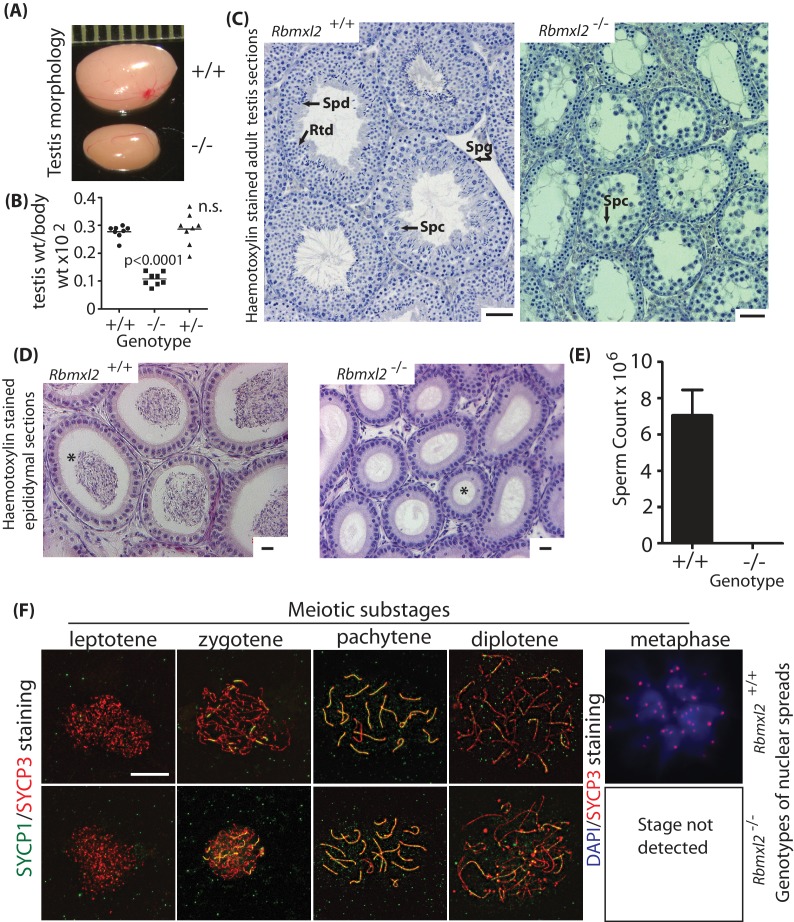
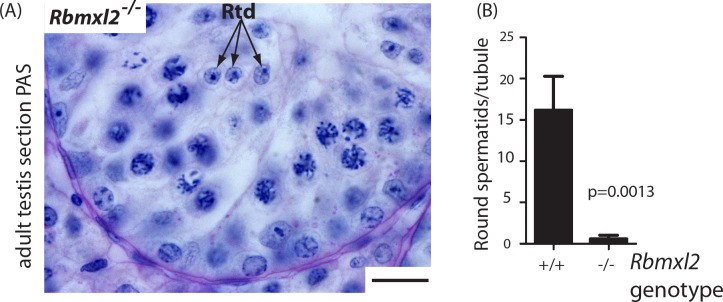
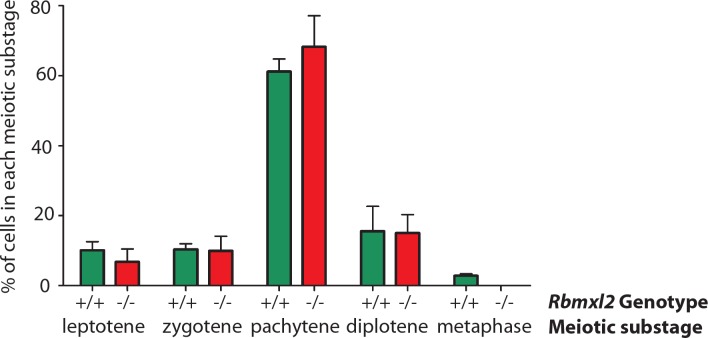
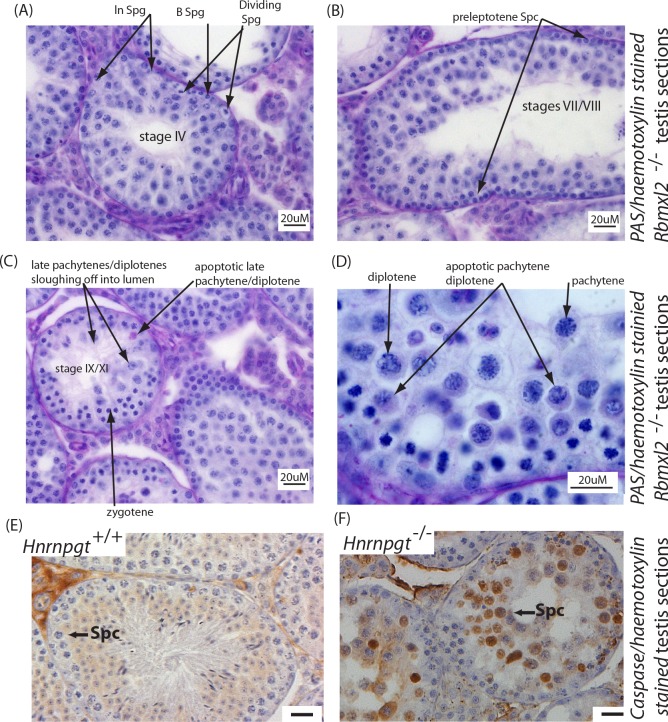
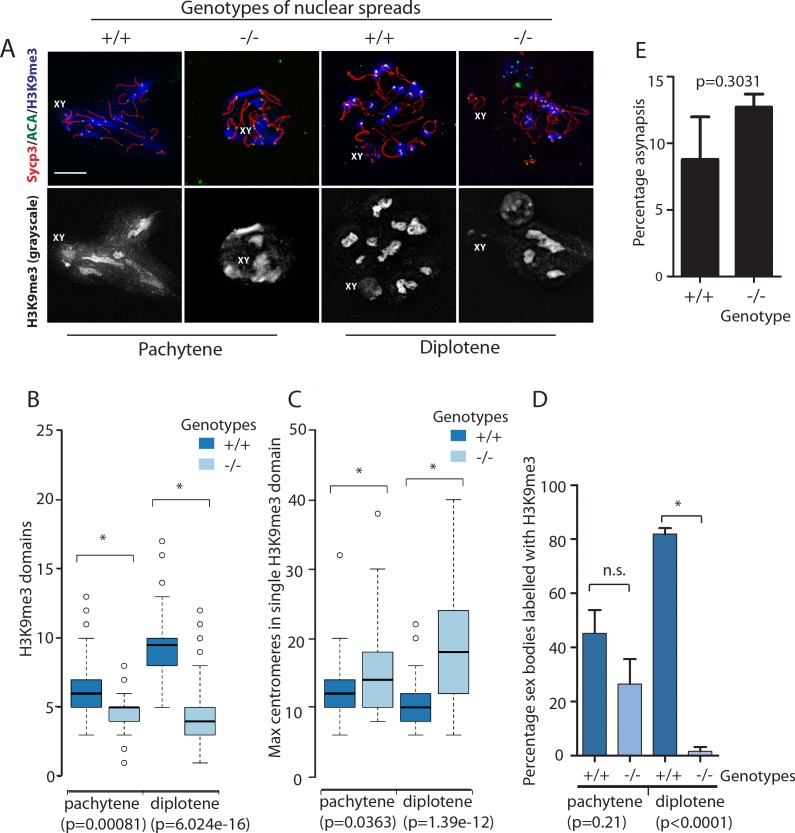
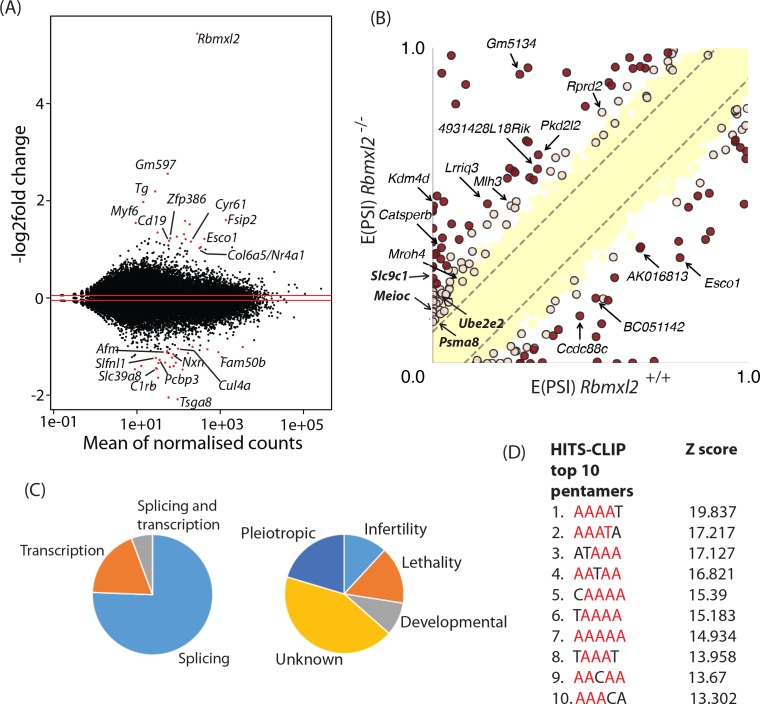
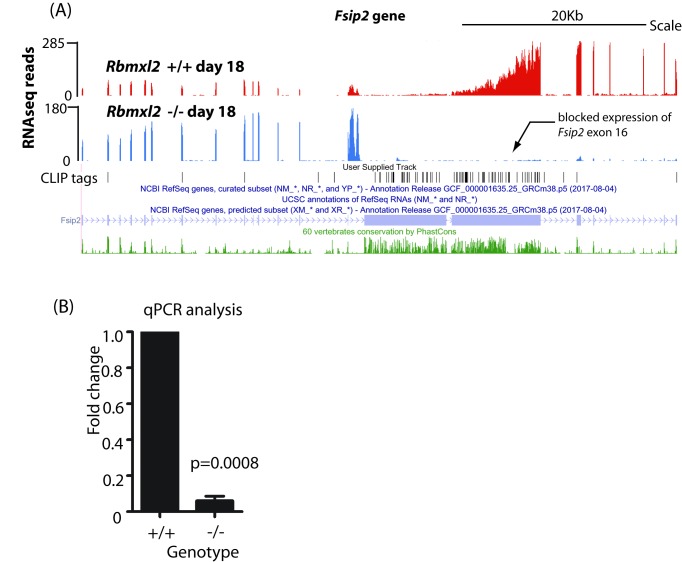
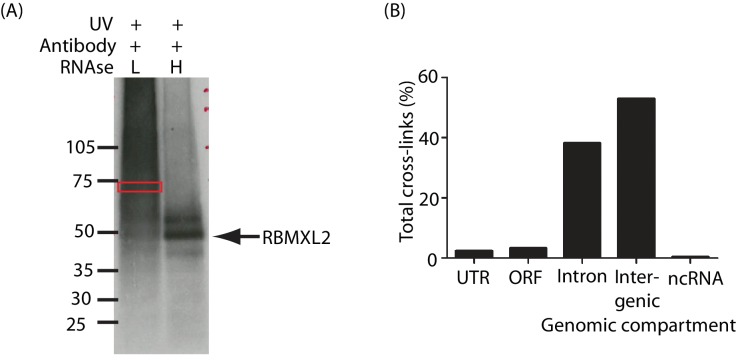
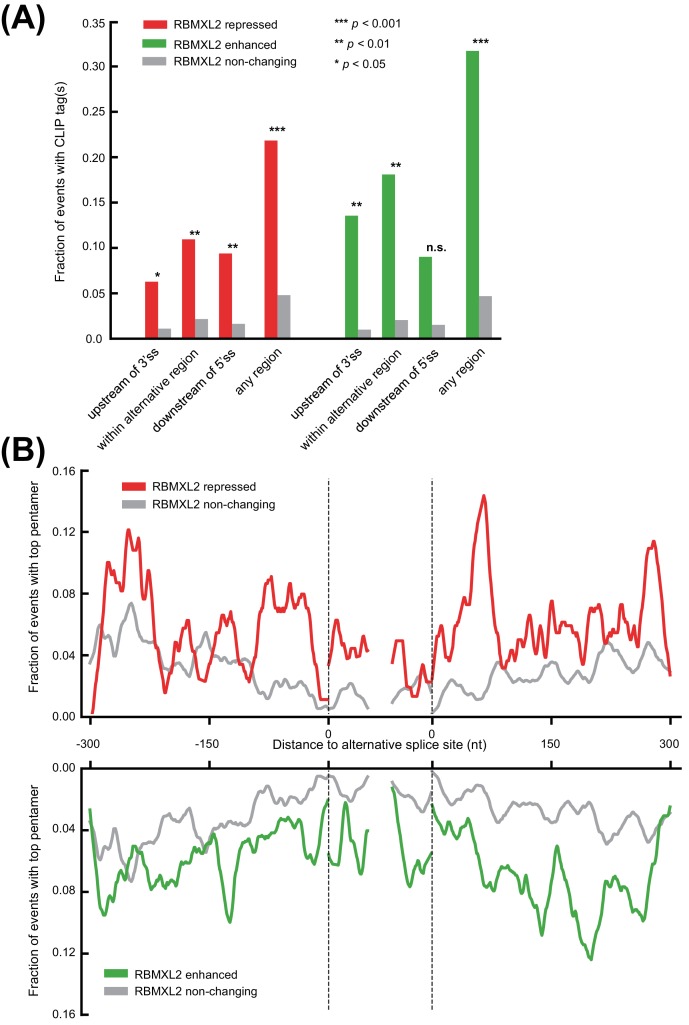
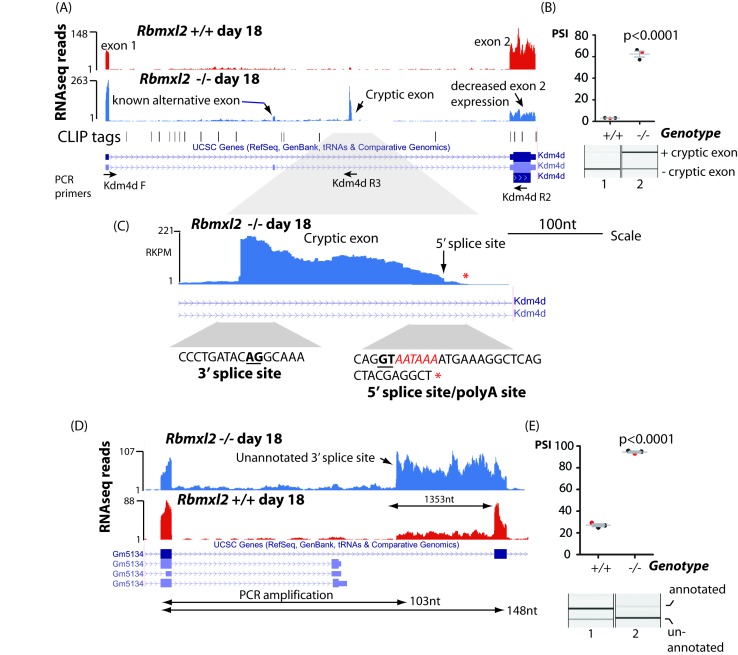
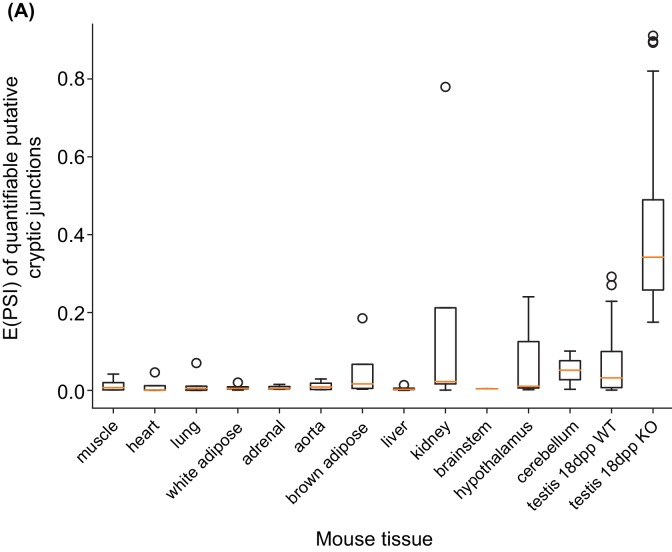
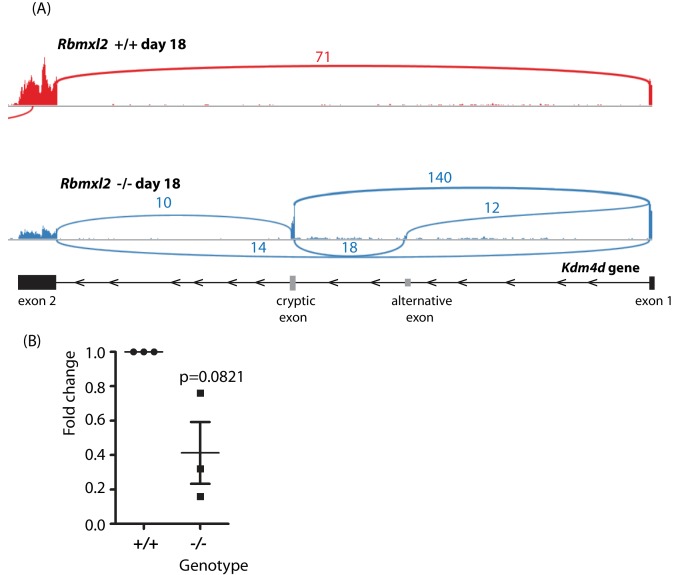
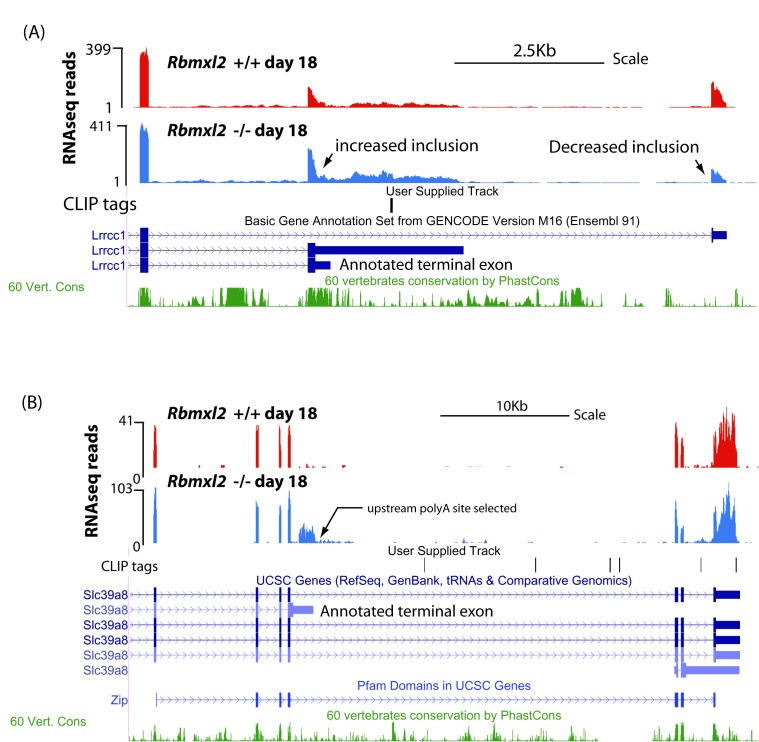
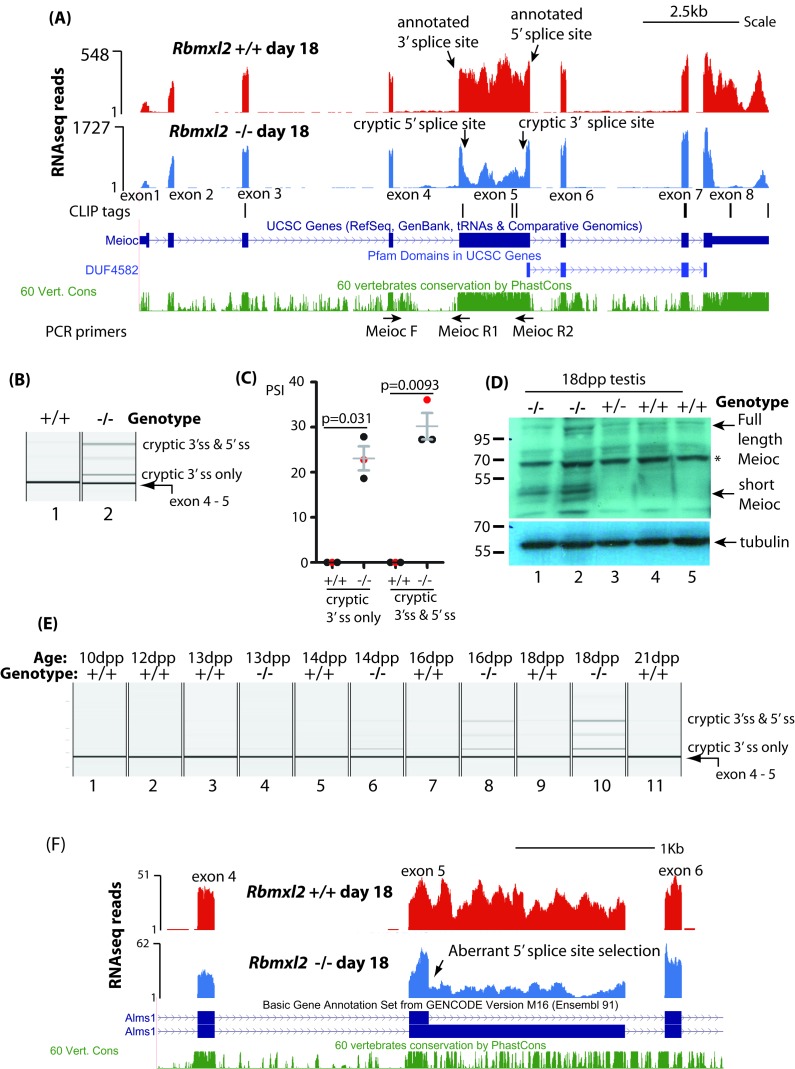
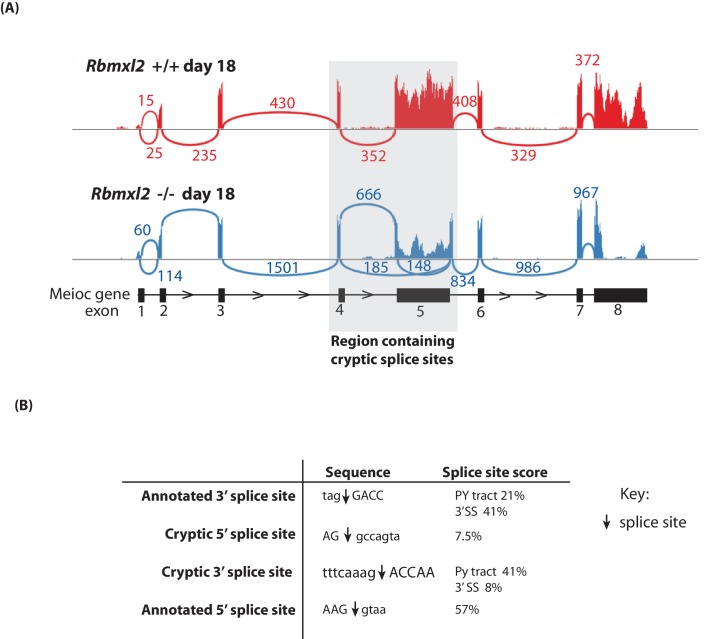
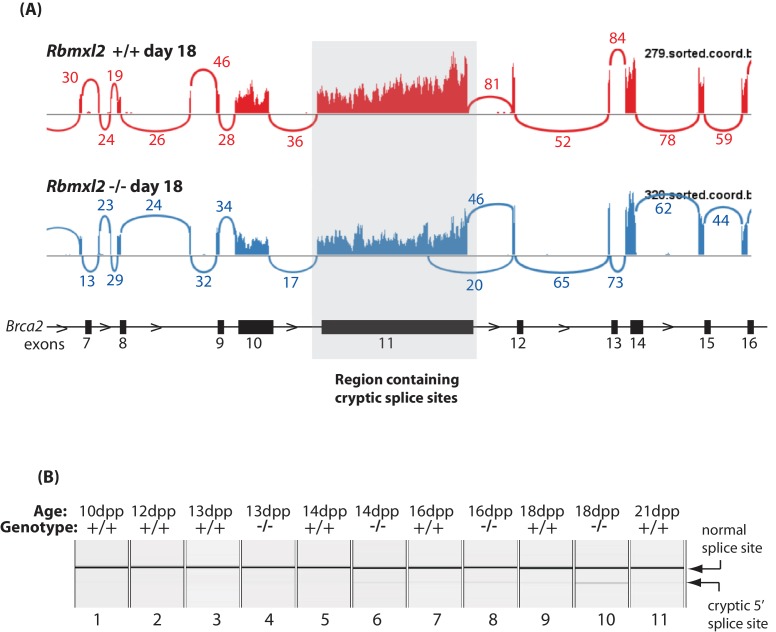
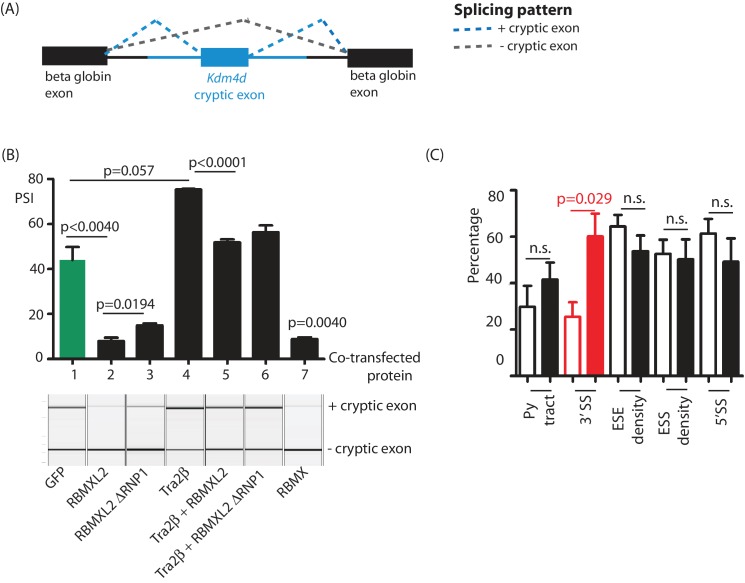
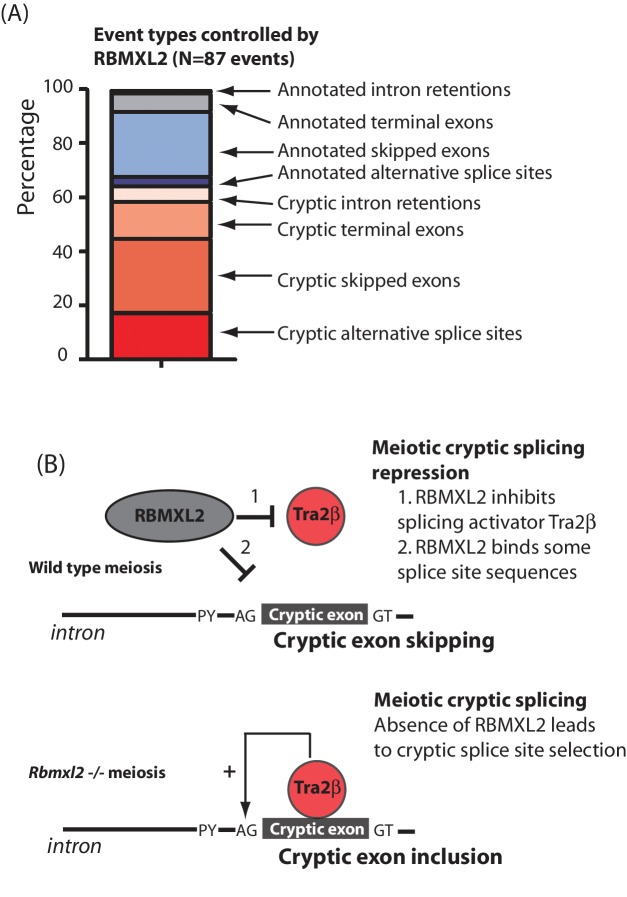
Male germ cells of all placental mammals express an ancient nuclear RNA binding protein of unknown function called RBMXL2. Here we find that deletion of the retrogene encoding RBMXL2 blocks spermatogenesis. Transcriptome analyses of age-matched deletion mice show that RBMXL2 controls splicing patterns during meiosis. In particular, RBMXL2 represses the selection of aberrant splice sites and the insertion of cryptic and premature terminal exons. Our data suggest a Rbmxl2 retrogene has been conserved across mammals as part of a splicing control mechanism that is fundamentally important to germ cell biology. We propose that this mechanism is essential to meiosis because it buffers the high ambient concentrations of splicing activators, thereby preventing poisoning of key transcripts and disruption to gene expression by aberrant splice site selection.
Keywords: RNA; alternative splicing; chromosomes; gene expression; meiosis; mouse; retrogene; spermatogenesis.
© 2019, Ehrmann et al.
Conflict of interest statement
IE, JC, MG, KJ, YL, SG, TC, Dd, JS, SC, IA, YB, DE No competing interests declared
Figures

References
-
- Abby E, Tourpin S, Ribeiro J, Daniel K, Messiaen S, Moison D, Guerquin J, Gaillard JC, Armengaud J, Langa F, Toth A, Martini E, Livera G. Implementation of meiosis prophase I programme requires a conserved retinoid-independent stabilizer of meiotic transcripts. Nature Communications. 2016;7:10324. doi: 10.1038/ncomms10324. - DOI - PMC - PubMed
-
- Arsov T, Silva DG, O'Bryan MK, Sainsbury A, Lee NJ, Kennedy C, Manji SS, Nelms K, Liu C, Vinuesa CG, de Kretser DM, Goodnow CC, Petrovsky N. Fat aussie--a new Alström syndrome mouse showing a critical role for ALMS1 in obesity, diabetes, and spermatogenesis. Molecular Endocrinology. 2006;20:1610–1622. doi: 10.1210/me.2005-0494. - DOI - PubMed
