

Lysyl oxidase-like 2 (LOXL2)-mediated cross-linking of tropoelastin
- PMID: 30676771
- PMCID: PMC6629125
- DOI: 10.1096/fj.201801860RR
Lysyl oxidase-like 2 (LOXL2)-mediated cross-linking of tropoelastin
Abstract
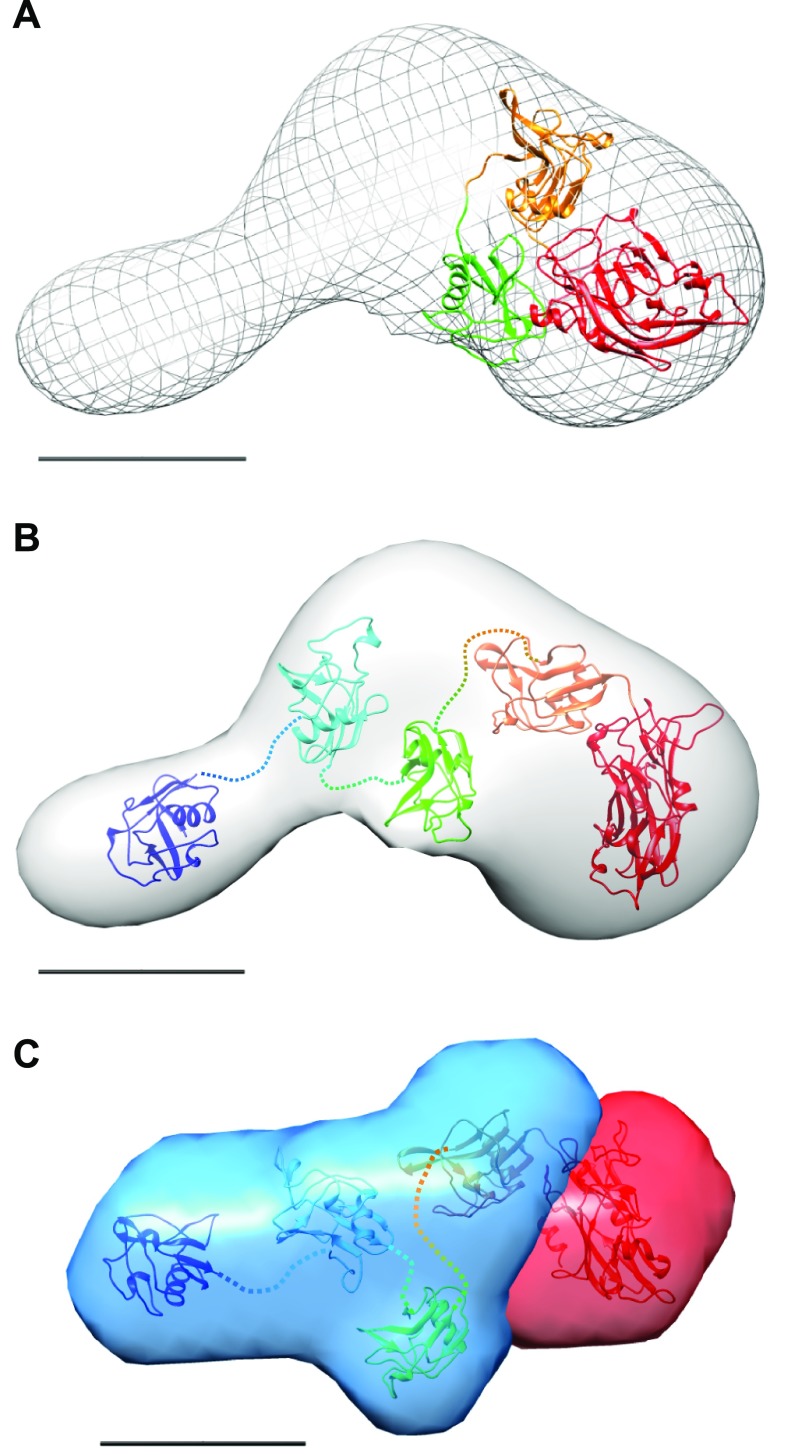
Lysyl oxidases (LOXs) play a central role in extracellular matrix remodeling during development and tumor growth and fibrosis through cross-linking of collagens and elastin. We have limited knowledge of the structure and substrate specificity of these secreted enzymes. LOXs share a conserved C-terminal catalytic domain but differ in their N-terminal region, which is composed of 4 repeats of scavenger receptor cysteine-rich (SRCR) domains in LOX-like (LOXL) 2. We investigated by X-ray scattering and electron microscopy the low-resolution structure of the full-length enzyme and the structure of a shorter form lacking the catalytic domain. Our data demonstrate that LOXL2 has a rod-like structure with a stalk composed of the SRCR domains and the catalytic domain at its tip. We detected direct interaction between LOXL2 and tropoelastin (TE) and also LOXL2-mediated deamination of TE. Using proteomics, we identified several allysines together with cross-linked TE peptides. The elastin-like material generated was resistant to trypsin proteolysis and displayed mechanical properties similar to mature elastin. Finally, we detected the codistribution of LOXL2 and elastin in the vascular wall. Altogether, these data suggest that LOXL2 could participate in elastogenesis in vivo and could be used as a means of cross-linking TE in vitro for biomimetic and cell-compatible tissue engineering purposes.-Schmelzer, C. E. H., Heinz, A., Troilo, H., Lockhart-Cairns, M.-P., Jowitt, T. A., Marchand, M. F., Bidault, L., Bignon, M., Hedtke, T., Barret, A., McConnell, J. C., Sherratt, M. J., Germain, S., Hulmes, D. J. S., Baldock, C., Muller, L. Lysyl oxidase-like 2 (LOXL2)-mediated cross-linking of tropoelastin.
Keywords: SAXS; elastin; matrix remodeling; protein structure; proteomics.
Conflict of interest statement
The authors thank the Structure Fédérative de Recherche Biosciences (UMS3444/CNRS), US8/INSERM, École Normale Supérieure de Lyon, Université Claude Bernard Lyon 1 Protein Science Facility for access to the circular dichroism instrument. The authors thank Christian Ihling (Martin Luther University Halle-Wittenberg, Germany) for assistance with mass spectrometry measurements, Prof. Tony Weiss (University of Sydney, Sydney, NSW, Australia) for providing the recombinant tropoelastin used in surface plasmon resonance experiments, Markus Pietzsch (Martin Luther University Halle-Wittenberg) for providing the recombinant tropoelastin used in other experiments, and Jürgen Brinckmann (University of Lübeck, Germany) for the amino acid analysis, and Dr. Catherine Moali (Université de Lyon, France) for helpful discussions and reading the manuscript, as well as providing BMP-1 and PCPE-1. The authors thank staff in the University of Manchester Biomolecular Analysis Core Facility and Electron Microscopy Centre for assistance and Diamond Light Source and European Synchrotron Radiation Facility (ESRF) (BioStruct-X FP7-Infractrustures Project 283570 Midland BAG MX1783) for beamtime. The work was supported by the Fraunhofer Internal Programs (Attract Grant 069–608203 to C.E.H.S.), the European Regional Development Fund in Saxony-Anhalt (project ElastoHEAL; to CEHS), the Biotechnology and Biological Sciences Research Council (BBSRC) (BB/L00612X/1 to C.B.), the LEO Foundation (Grant LF17063 to A.H.), and Prevent Breast Cancer (to M.J.S.). M.F.M. was supported by Fédération pour la Recherche Médicale (FRM-ECO2016-0736055). The authors declare no conflicts of interest.
Figures







References
-
- Trackman P. C., Bais M. V. (2018) Measurement of lysyl oxidase activity from small tissue samples and cell cultures. Methods Cell Biol. 143, 147–156 - PubMed
-
- Mäki J. M., Räsänen J., Tikkanen H., Sormunen R., Mäkikallio K., Kivirikko K. I., Soininen R. (2002) Inactivation of the lysyl oxidase gene Lox leads to aortic aneurysms, cardiovascular dysfunction, and perinatal death in mice. Circulation 106, 2503–2509 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
