

Precision in the development of neocortical architecture: From progenitors to cortical networks
- PMID: 30677429
- PMCID: PMC6402587
- DOI: 10.1016/j.pneurobio.2019.01.003
Precision in the development of neocortical architecture: From progenitors to cortical networks
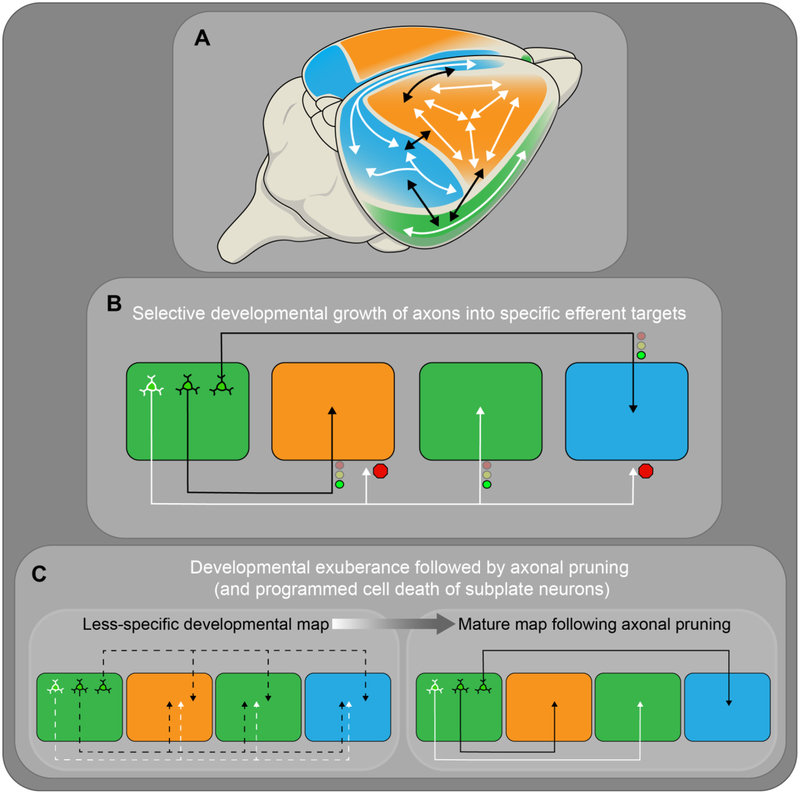
Abstract
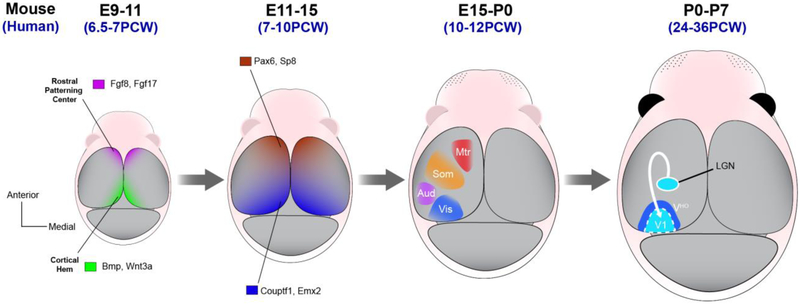
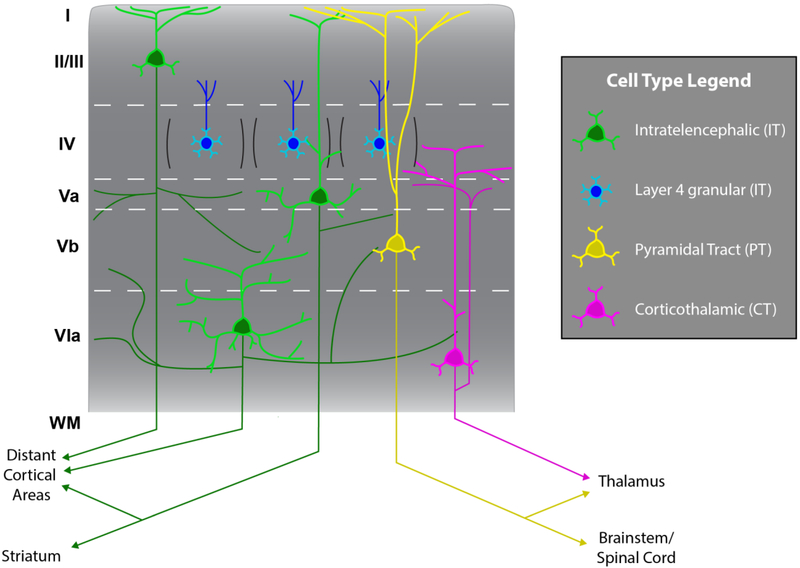
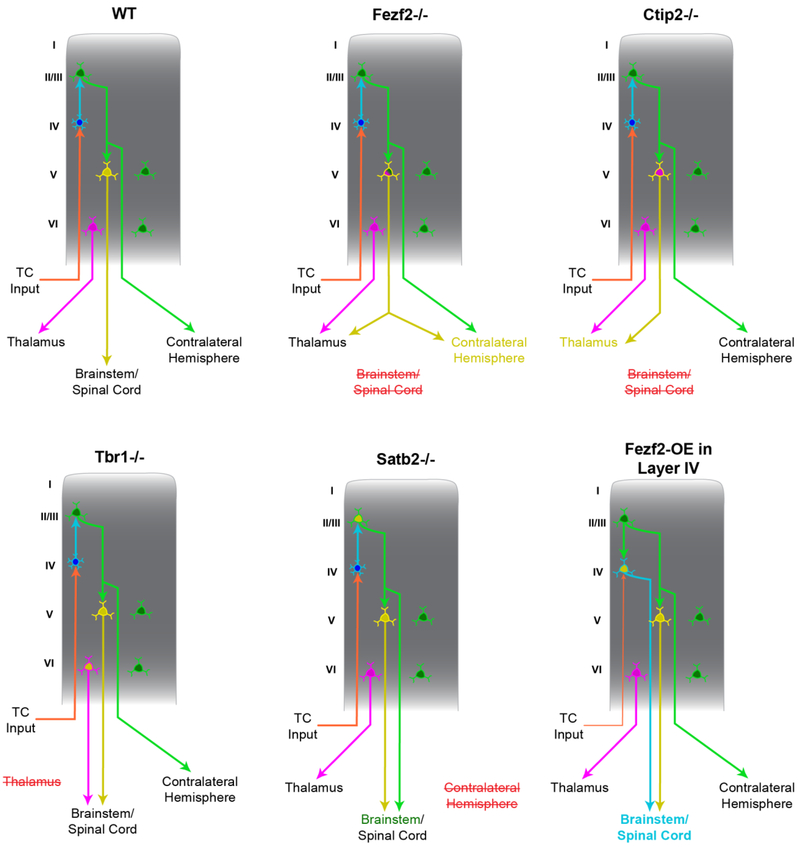
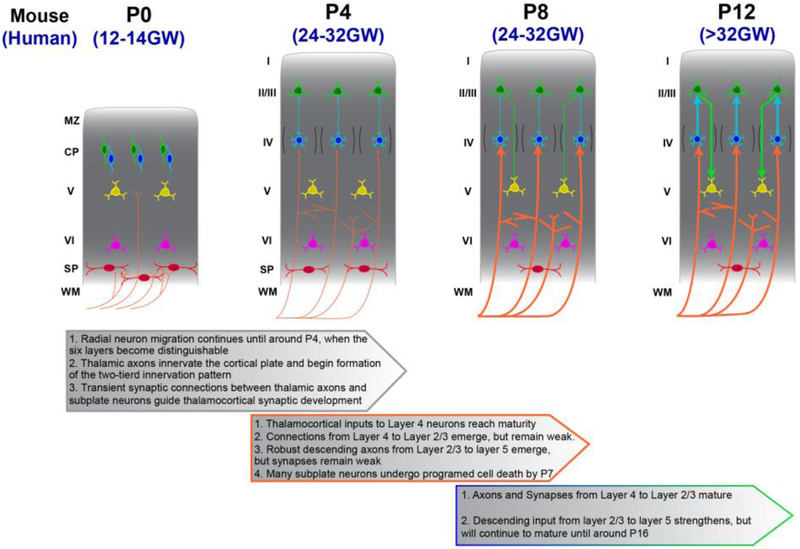
Of all brain regions, the 6-layered neocortex has undergone the most dramatic changes in size and complexity during mammalian brain evolution. These changes, occurring in the context of a conserved set of organizational features that emerge through stereotypical developmental processes, are considered responsible for the cognitive capacities and sensory specializations represented within the mammalian clade. The modern experimental era of developmental neurobiology, spanning 6 decades, has deciphered a number of mechanisms responsible for producing the diversity of cortical neuron types, their precise connectivity and the role of gene by environment interactions. Here, experiments providing insight into the development of cortical projection neuron differentiation and connectivity are reviewed. This current perspective integrates discussion of classic studies and new findings, based on recent technical advances, to highlight an improved understanding of the neuronal complexity and precise connectivity of cortical circuitry. These descriptive advances bring new opportunities for studies related to the developmental origins of cortical circuits that will, in turn, improve the prospects of identifying pathogenic targets of neurodevelopmental disorders.
Keywords: Axon guidance; Cell type specification; Cerebral cortex; Circuits; Connectivity; Excitatory neurons; Experience; Genetics; Histogenesis; Human; Microcircuit; Neural network; Refinement; Reprogramming; Rodent; Specification; Synaptic specificity; Synaptogenesis; Thalamus.
Copyright © 2019 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Alcamo EA, Chirivella L, Dautzenberg M, Dobreva G, Farin I, Grosschedl R, McConnell SK (2008) Satb2 Regulates Callosal Projection Neuron Identity in the Developing Cerebral Cortex. Neuron 57:364–377. - PubMed
-
- Angevine JJ, Sidman R (1961) Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse.. Nature:766–768. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
