

Deciphering the Molecular Mechanisms Sustaining the Estrogenic Activity of the Two Major Dietary Compounds Zearalenone and Apigenin in ER-Positive Breast Cancer Cell Lines
- PMID: 30678243
- PMCID: PMC6412274
- DOI: 10.3390/nu11020237
Deciphering the Molecular Mechanisms Sustaining the Estrogenic Activity of the Two Major Dietary Compounds Zearalenone and Apigenin in ER-Positive Breast Cancer Cell Lines
Abstract
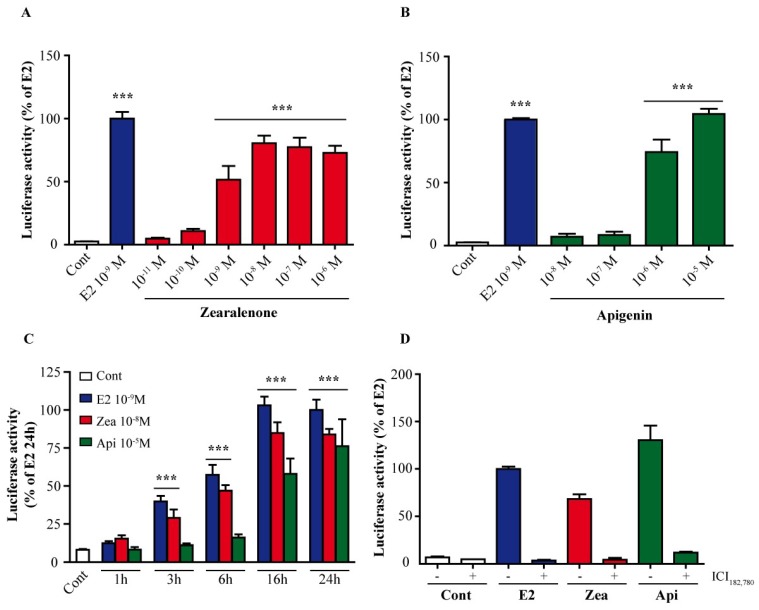
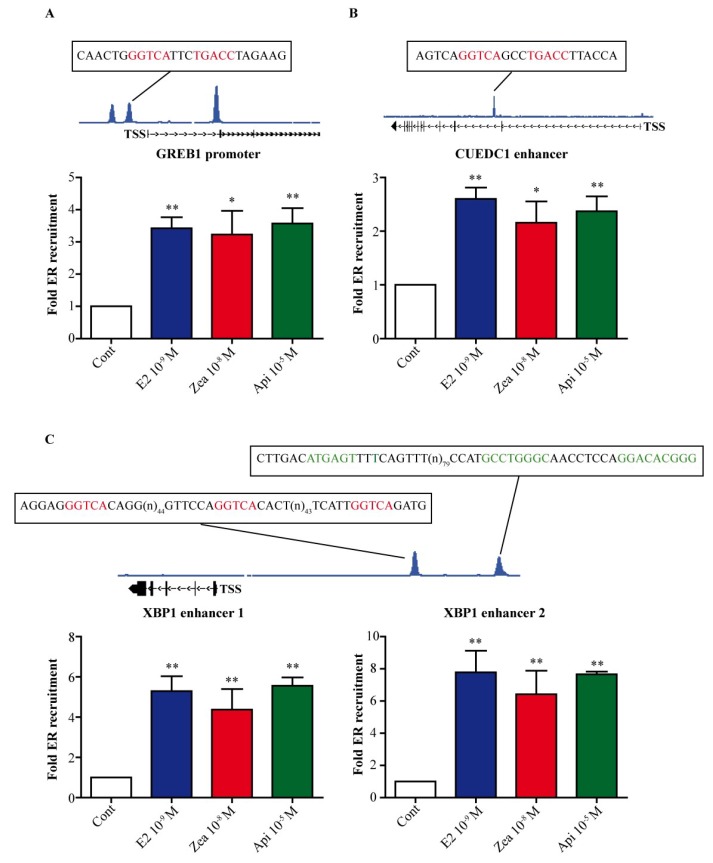
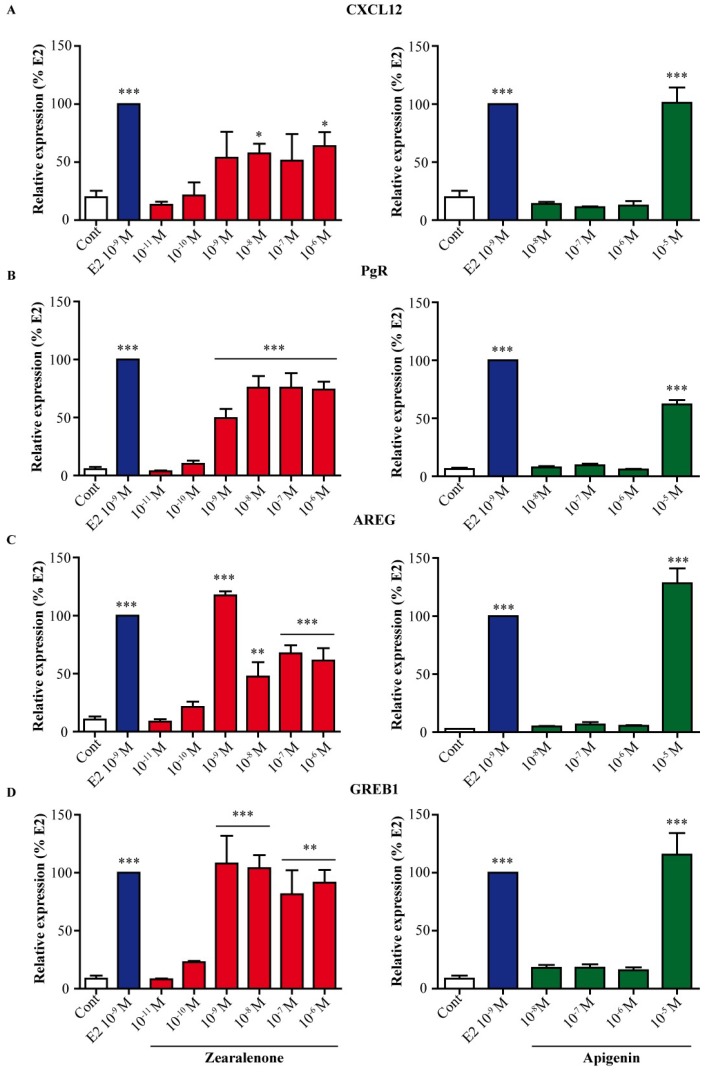
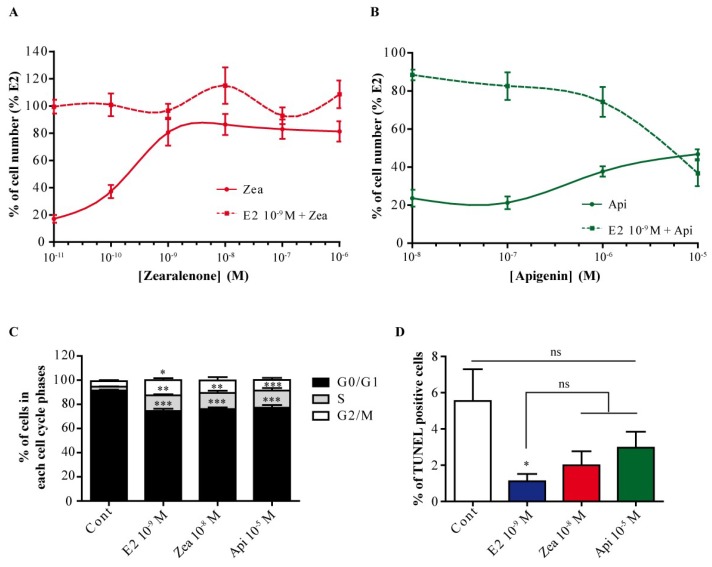
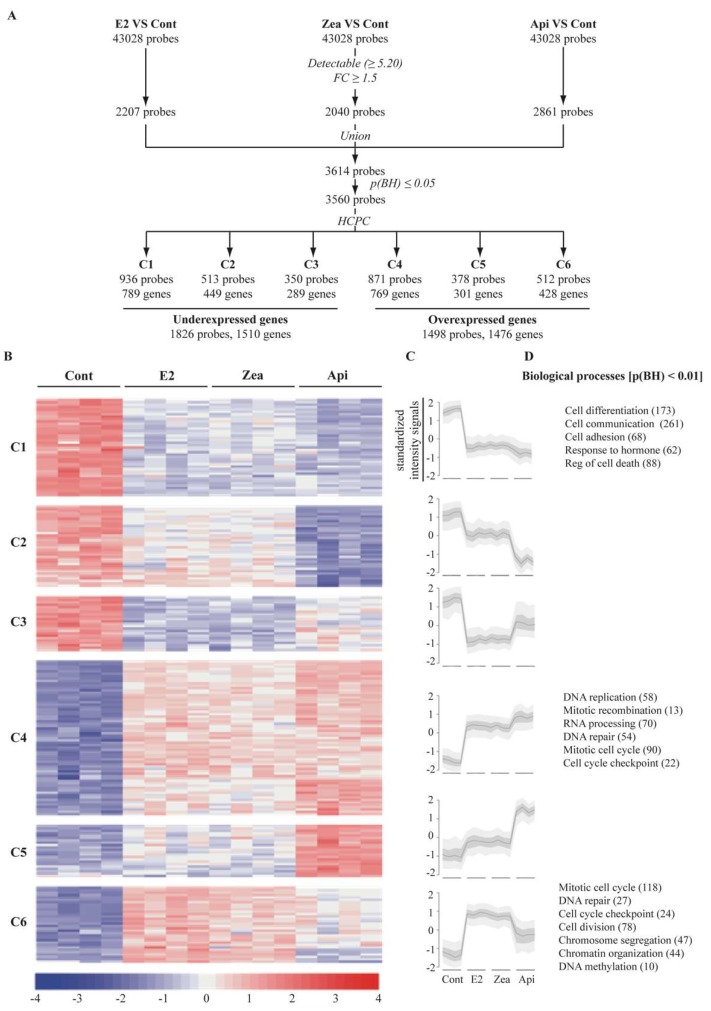
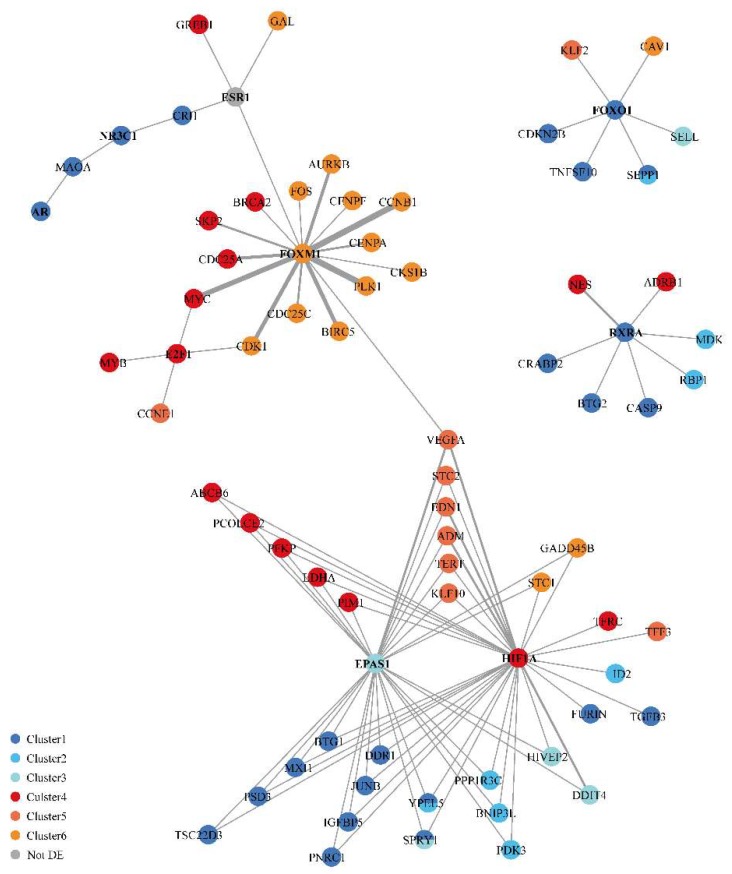
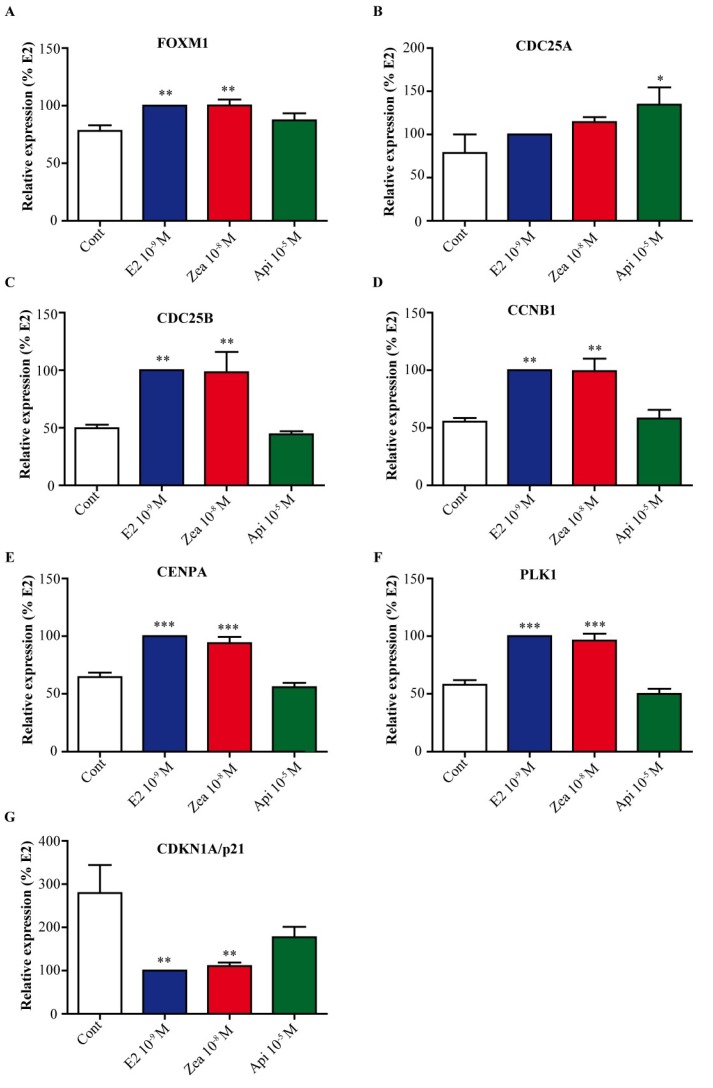
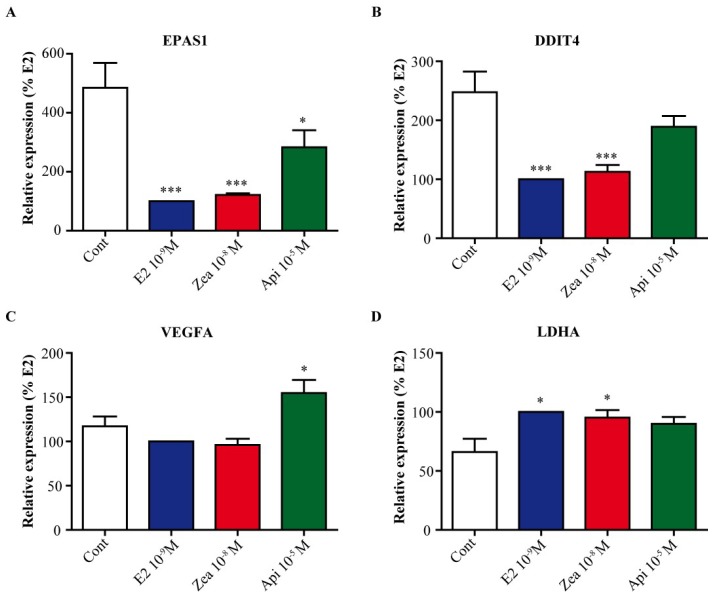
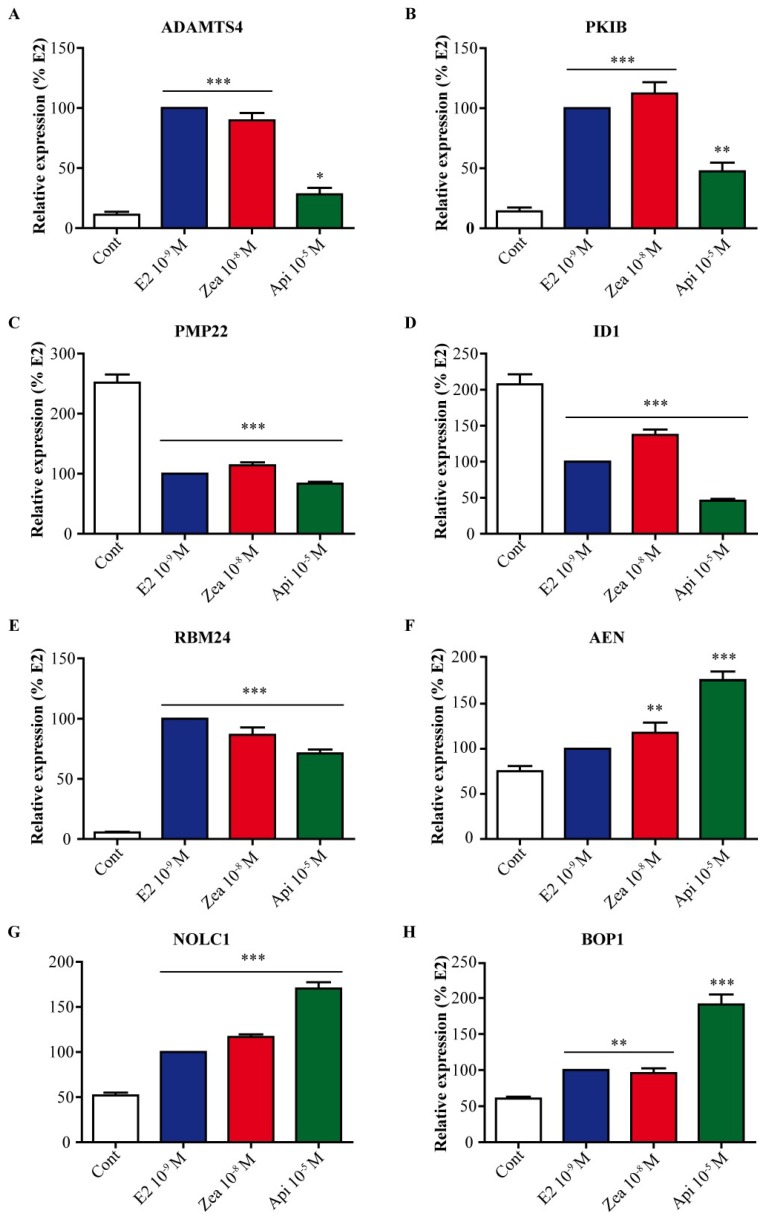
The flavone apigenin and the mycotoxin zearalenone are two major compounds found in the human diet which bind estrogen receptors (ERs), and therefore influence ER activity. However, the underlying mechanisms are not well known. To unravel the molecular mechanisms that could explain the differential effect of zearalenone and apigenin on ER-positive breast cancer cell proliferation, gene-reporter assays, chromatin immunoprecipitation (ChIP) experiments, proliferation assays and transcriptomic analysis were performed. We found that zearalenone and apigenin transactivated ERs and promoted the expression of estradiol (E2)-responsive genes. However, zearalenone clearly enhanced cellular proliferation, while apigenin appeared to be antiestrogenic in the presence of E2 in both ER-positive breast cancer cell lines, MCF-7 and T47D. The transcriptomic analysis showed that both compounds regulate gene expression in the same way, but with differences in intensity. Two major sets of genes were identified; one set was linked to the cell cycle and the other set was linked to stress response and growth arrest. Our results show that the transcription dynamics in gene regulation induced by apigenin were somehow different with zearalenone and E2 and may explain the differential effect of these compounds on the phenotype of the breast cancer cell. Together, our results confirmed the potential health benefit effect of apigenin, while zearalenone appeared to be a true endocrine-disrupting compound.
Keywords: breast cancer; dietary compounds; endocrine disruption; estrogen receptor; gene expression.
Conflict of interest statement
The authors declare they have no actual or potential competing financial interests.
Figures
References
-
- Crain D.A., Janssen S.J., Edwards T.M., Heindel J., Ho S., Hunt P., Iguchi T., Juul A., McLachlan J.A., Schwartz J., et al. Female reproductive disorders: The roles of endocrine-disrupting compounds and developmental timing. Fertil. Steril. 2008;90:911–940. doi: 10.1016/j.fertnstert.2008.08.067. - DOI - PMC - PubMed
-
- Lecomte S., Lelong M., Bourgine G., Efstathiou T., Saligaut C., Pakdel F. Assessment of the potential activity of major dietary compounds as selective estrogen receptor modulators in two distinct cell models for proliferation and differentiation. Toxicol. Appl. Pharmacol. 2017;325:61–70. doi: 10.1016/j.taap.2017.04.005. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
