

T-Cell Response to Viral Hemorrhagic Fevers
- PMID: 30678246
- PMCID: PMC6466054
- DOI: 10.3390/vaccines7010011
T-Cell Response to Viral Hemorrhagic Fevers
Abstract
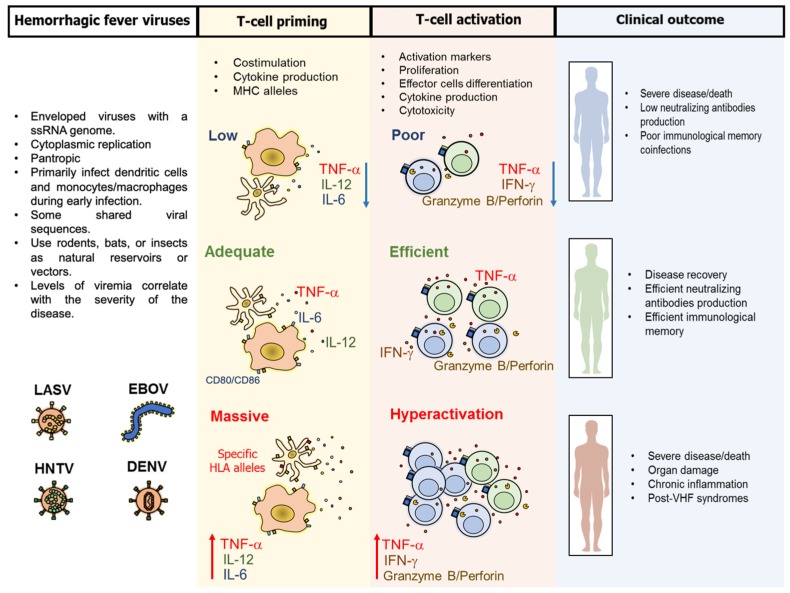
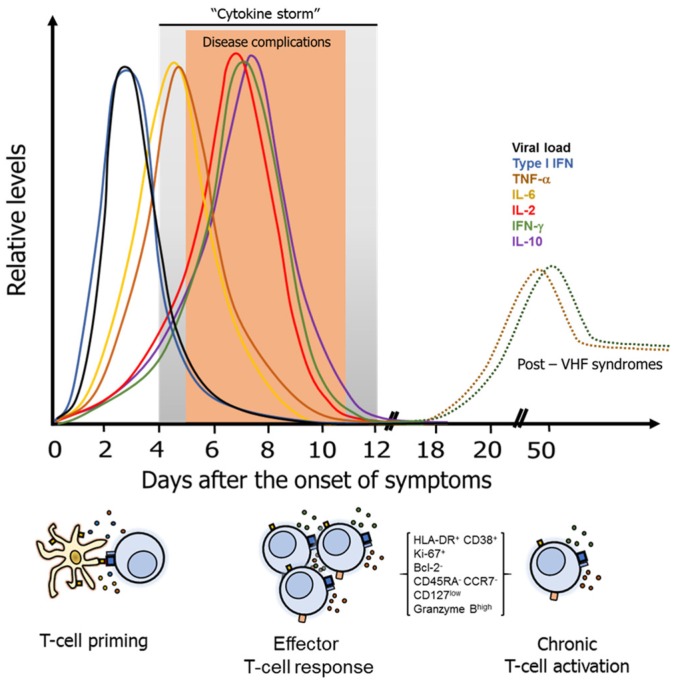
Viral hemorrhagic fevers (VHF) are a group of clinically similar diseases that can be caused by enveloped RNA viruses primarily from the families Arenaviridae, Filoviridae, Hantaviridae, and Flaviviridae. Clinically, this group of diseases has in common fever, fatigue, dizziness, muscle aches, and other associated symptoms that can progress to vascular leakage, bleeding and multi-organ failure. Most of these viruses are zoonotic causing asymptomatic infections in the primary host, but in human beings, the infection can be lethal. Clinical and experimental evidence suggest that the T-cell response is needed for protection against VHF, but can also cause damage to the host, and play an important role in disease pathogenesis. Here, we present a review of the T-cell immune responses to VHF and insights into the possible ways to improve counter-measures for these viral agents.
Keywords: Ebola virus; Lassa virus; T-cells; dengue virus; hantavirus; interferon-gamma; tumor necrosis factor-alpha; vaccine; viral hemorrhagic fever; yellow fever virus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Animal models of viral hemorrhagic fever.Antiviral Res. 2014 Dec;112:59-79. doi: 10.1016/j.antiviral.2014.10.001. Epub 2014 Oct 18. Antiviral Res. 2014. PMID: 25448088 Review.
-
Molecular pathogenesis of viral hemorrhagic fever.Semin Immunopathol. 2017 Jul;39(5):551-561. doi: 10.1007/s00281-017-0637-x. Epub 2017 May 29. Semin Immunopathol. 2017. PMID: 28555386 Free PMC article. Review.
-
Hemorrhagic fever virus-induced changes in hemostasis and vascular biology.Blood Coagul Fibrinolysis. 2000 Jul;11(5):461-83. doi: 10.1097/00001721-200007000-00010. Blood Coagul Fibrinolysis. 2000. PMID: 10937808 Review.
-
Immunological features underlying viral hemorrhagic fevers.Curr Opin Immunol. 2015 Oct;36:38-46. doi: 10.1016/j.coi.2015.06.003. Epub 2015 Jul 7. Curr Opin Immunol. 2015. PMID: 26163194 Free PMC article. Review.
-
Factors affecting RIG-I-Like receptors activation - New research direction for viral hemorrhagic fevers.Front Immunol. 2022 Sep 29;13:1010635. doi: 10.3389/fimmu.2022.1010635. eCollection 2022. Front Immunol. 2022. PMID: 36248895 Free PMC article. Review.
Cited by
-
Comparative Immunoreactivity Analyses of Hantaan Virus Glycoprotein-Derived MHC-I Epitopes in Vaccination.Vaccines (Basel). 2022 Apr 6;10(4):564. doi: 10.3390/vaccines10040564. Vaccines (Basel). 2022. PMID: 35455313 Free PMC article.
-
Low Activation of CD8+ T Cells in response to Viral Peptides in Mexican Patients with Severe Dengue.J Immunol Res. 2022 Mar 25;2022:9967594. doi: 10.1155/2022/9967594. eCollection 2022. J Immunol Res. 2022. PMID: 35372587 Free PMC article.
-
A Single Dose of Modified Vaccinia Ankara Expressing Lassa Virus-like Particles Protects Mice from Lethal Intra-cerebral Virus Challenge.Pathogens. 2019 Aug 28;8(3):133. doi: 10.3390/pathogens8030133. Pathogens. 2019. PMID: 31466243 Free PMC article.
-
Vascular dysfunction in hemorrhagic viral fevers: opportunities for organotypic modeling.Biofabrication. 2024 Jun 5;16(3):032008. doi: 10.1088/1758-5090/ad4c0b. Biofabrication. 2024. PMID: 38749416 Free PMC article. Review.
-
Potent immunogenicity and protective efficacy of a multi-pathogen vaccination targeting Ebola, Sudan, Marburg and Lassa viruse.PLoS Pathog. 2024 Jun 26;20(6):e1012262. doi: 10.1371/journal.ppat.1012262. eCollection 2024 Jun. PLoS Pathog. 2024. PMID: 38924060 Free PMC article.
References
-
- Salvato M.S. Hemorrhagic Fever Viruses. 1st ed. Springer Nature; New York, NY, USA: 2018.
Publication types
LinkOut - more resources
Full Text Sources
