

The Striatum's Role in Executing Rational and Irrational Economic Behaviors
- PMID: 30678530
- PMCID: PMC6656632
- DOI: 10.1177/1073858418824256
The Striatum's Role in Executing Rational and Irrational Economic Behaviors
Abstract
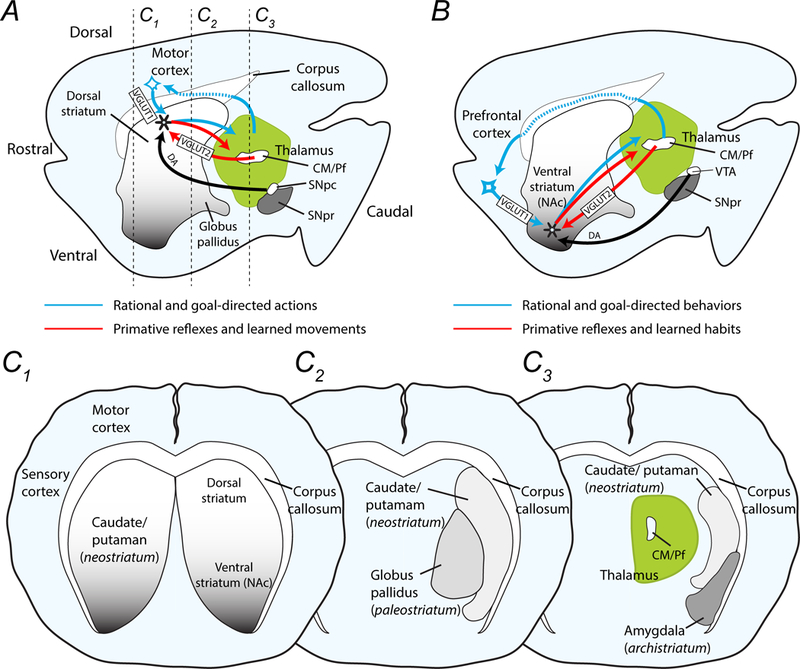
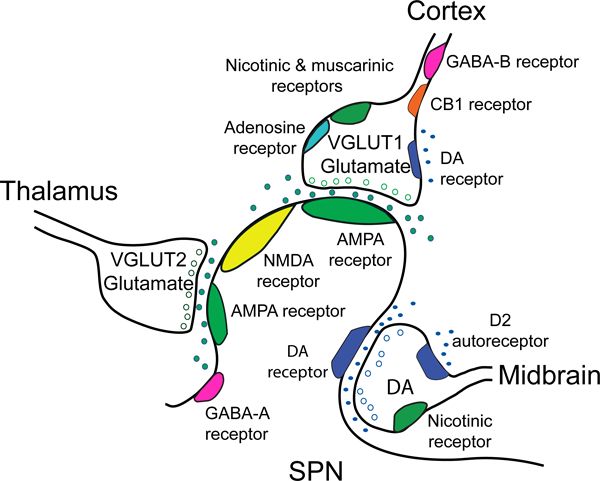
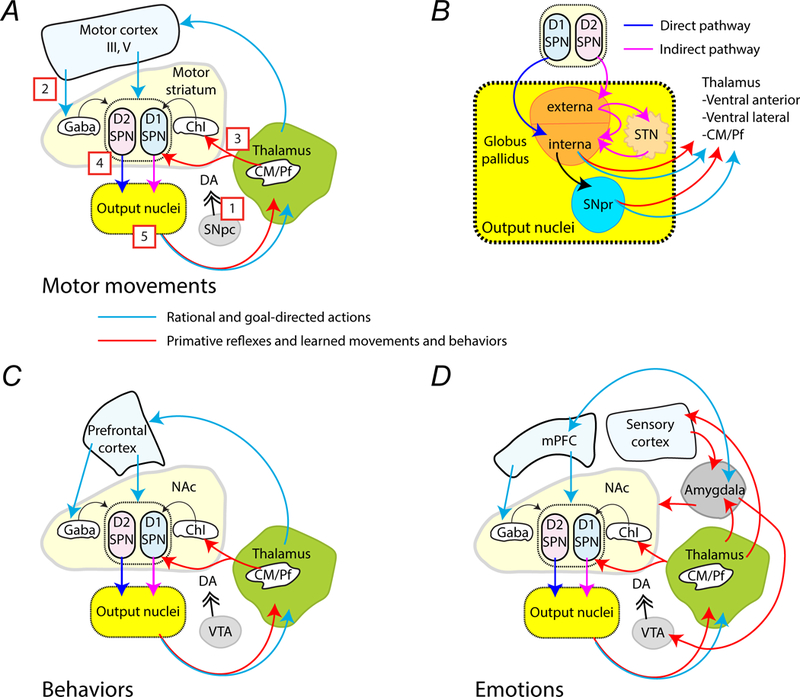
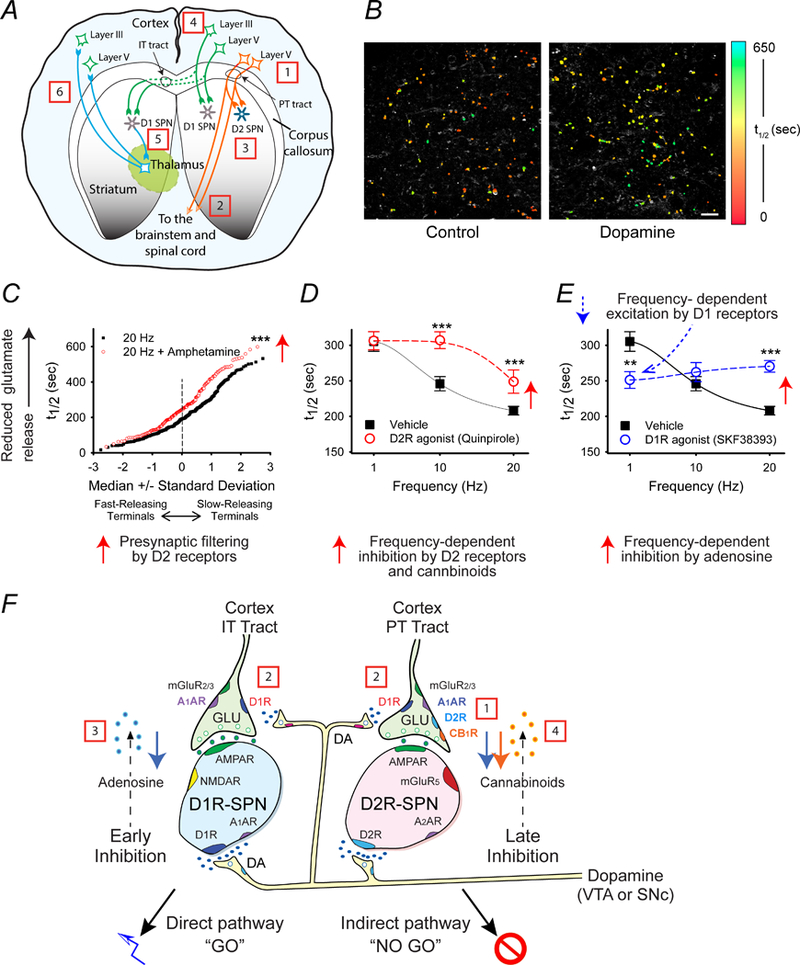
The striatum is a critical component of the brain that controls motor, reward, and executive function. This ancient and phylogenetically conserved structure forms a central hub where rapid instinctive, reflexive movements and behaviors in response to sensory stimulation or the retrieval of emotional memory intersect with slower planned motor movements and rational behaviors. This review emphasizes two distinct pathways that begin in the thalamus and converge in the striatum to differentially affect movements, behaviors, and decision making. The convergence of excitatory glutamatergic activity from the thalamus and cortex, along with dopamine release in response to novel stimulation, provide the basis for motor learning, reward seeking, and habit formation. We outline how the rules derived through research on neural pathways may enhance the predictability of reflexive actions and rational responses studied in behavioral economics.
Keywords: behavioral economics; decision-making; emotion; habits; motor learning; neuroeconomics; neuroscience; reward.
Conflict of interest statement
Declaration of Conflicting Interests
The authors declare that there is no conflict of interest
Figures
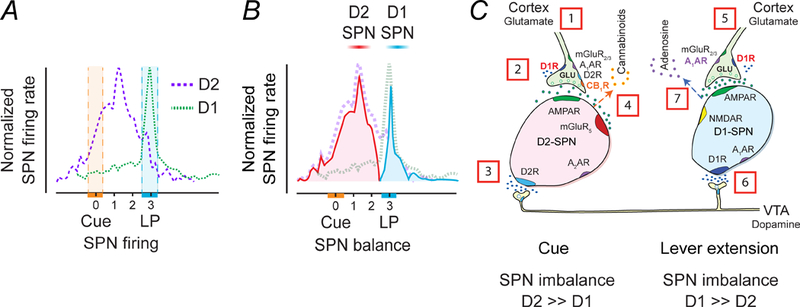
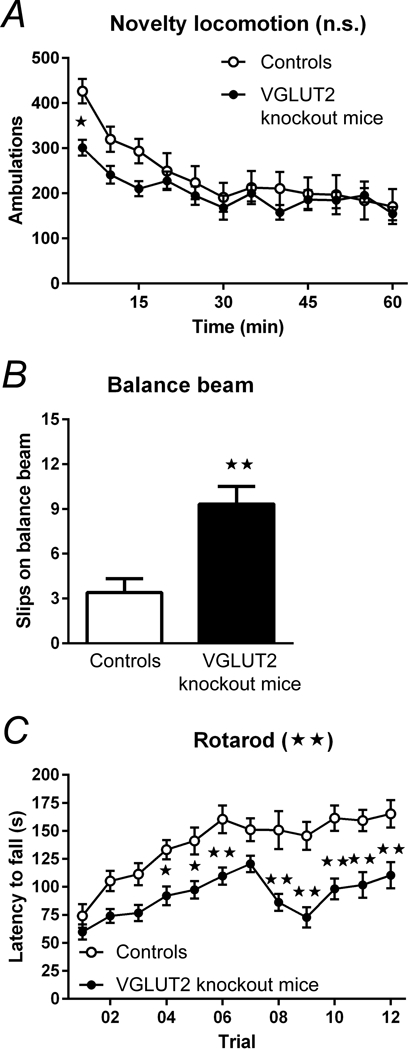
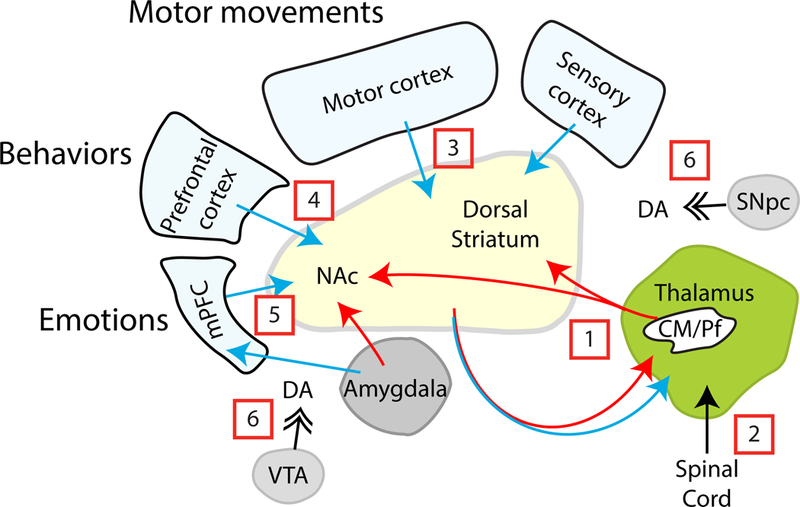
References
-
- Bamford NS, Joyce JA. 2005. Chronic methamphetamine mediates long-term depression of corticostriatal release in the dorsal striatum. Annals of Neurology 58(S9):S81.
-
- Bamford NS, Zhang H, Schmitz Y, Wu NP, Cepeda C, Levine MS and others. 2004b. Heterosynaptic dopamine neurotransmission selects sets of corticostriatal terminals. Neuron 42(4):653–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
