

Prominent action of butyrate over β-hydroxybutyrate as histone deacetylase inhibitor, transcriptional modulator and anti-inflammatory molecule
- PMID: 30679586
- PMCID: PMC6346118
- DOI: 10.1038/s41598-018-36941-9
Prominent action of butyrate over β-hydroxybutyrate as histone deacetylase inhibitor, transcriptional modulator and anti-inflammatory molecule
Abstract
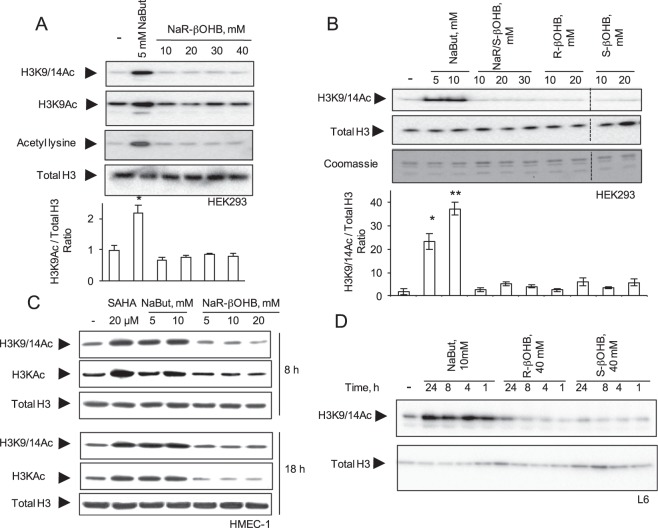
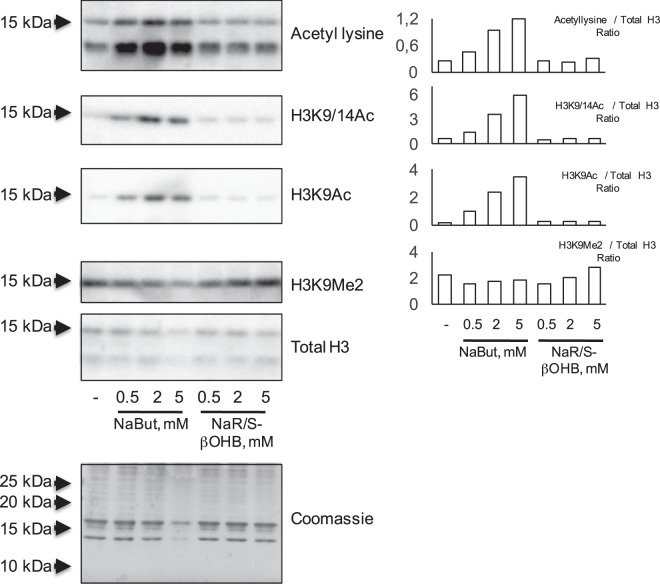
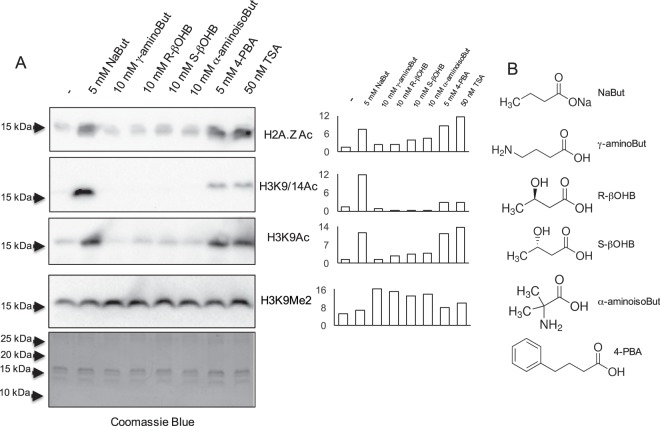
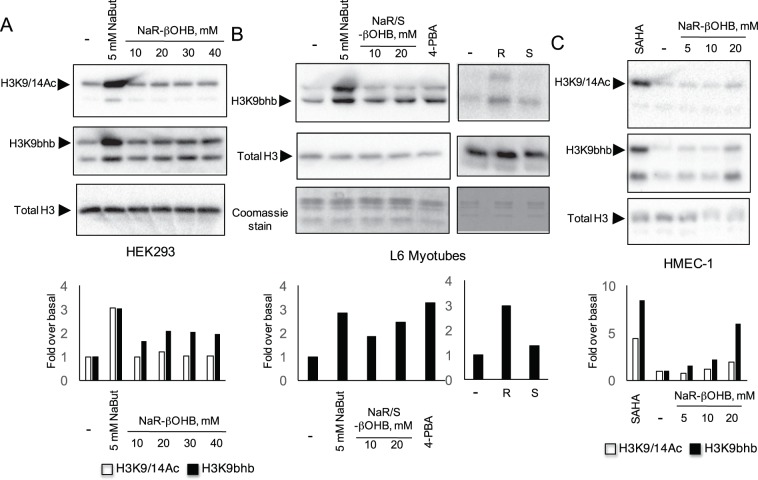
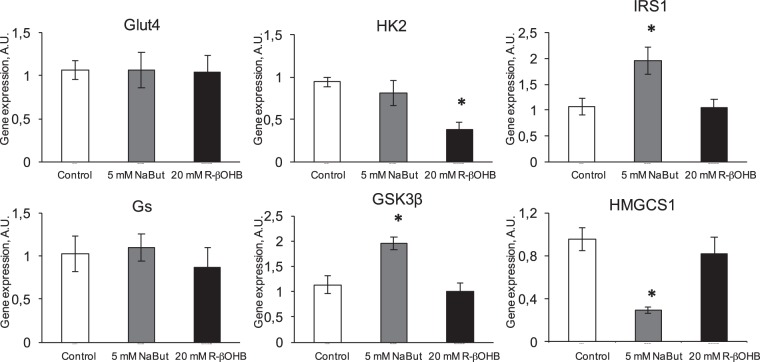
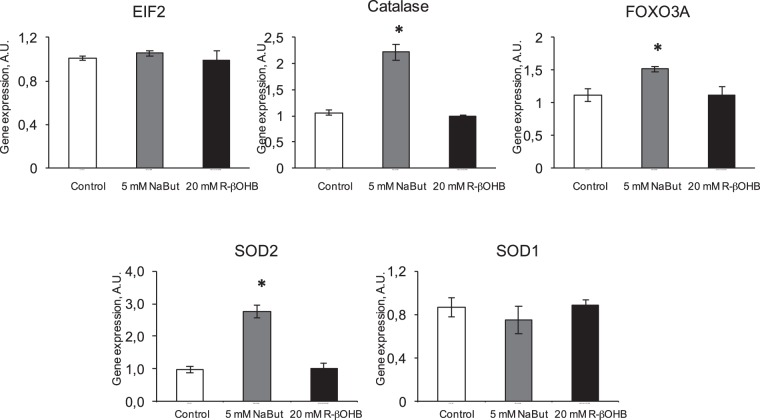
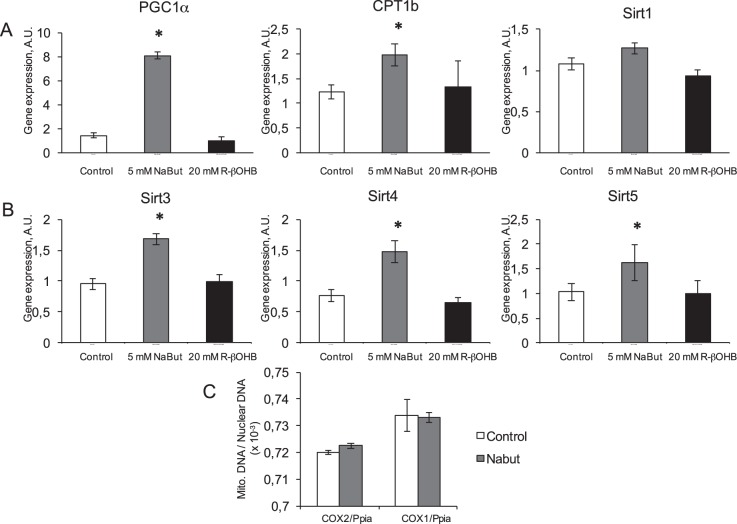
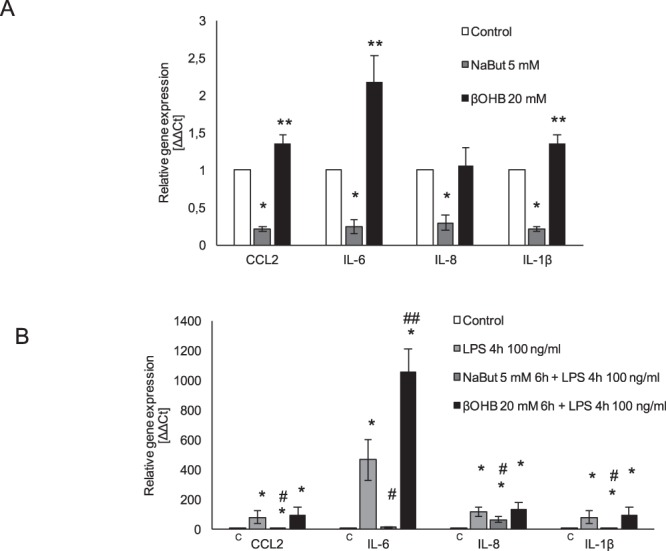
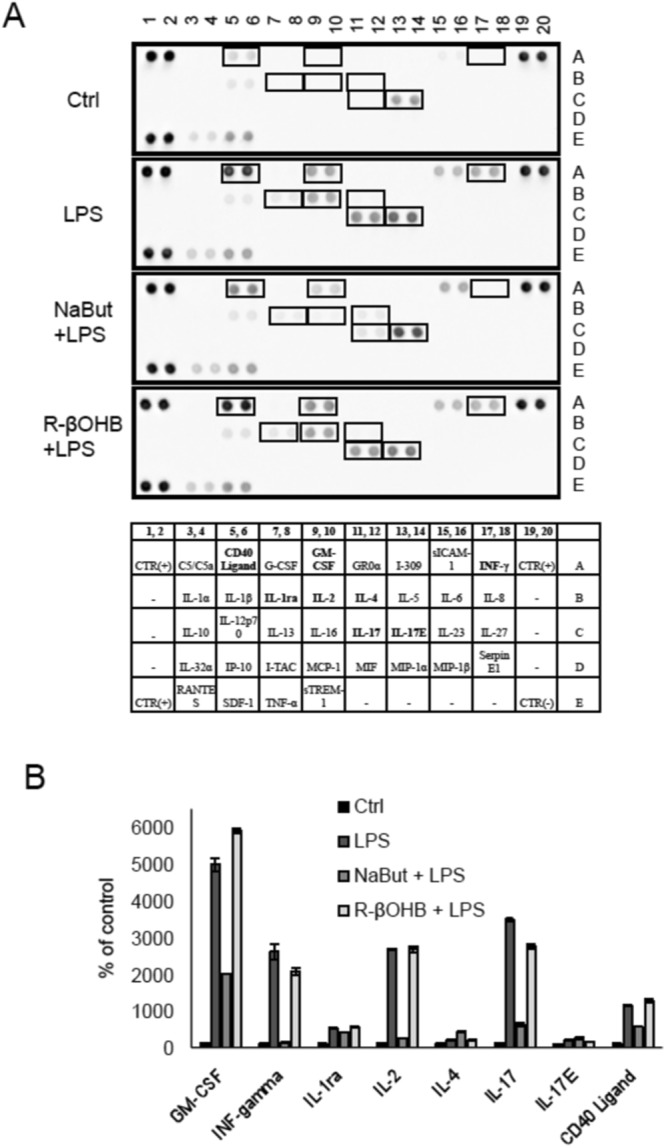
Butyrate and R-β-hydroxybutyrate are two related short chain fatty acids naturally found in mammals. Butyrate, produced by enteric butyric bacteria, is present at millimolar concentrations in the gastrointestinal tract and at lower levels in blood; R-β-hydroxybutyrate, the main ketone body, produced by the liver during fasting can reach millimolar concentrations in the circulation. Both molecules have been shown to be histone deacetylase (HDAC) inhibitors, and their administration has been associated to an improved metabolic profile and better cellular oxidative status, with butyrate inducing PGC1α and fatty acid oxidation and R-β-hydroxybutyrate upregulating oxidative stress resistance factors FOXO3A and MT2 in mouse kidney. Because of the chemical and functional similarity between the two molecules, we compared here their impact on multiple cell types, evaluating i) histone acetylation and hydroxybutyrylation levels by immunoblotting, ii) transcriptional regulation of metabolic and inflammatory genes by quantitative PCR and iii) cytokine secretion profiles using proteome profiling array analysis. We confirm that butyrate is a strong HDAC inhibitor, a characteristic we could not identify in R-β-hydroxybutyrate in vivo nor in vitro. Butyrate had an extensive impact on gene transcription in rat myotubes, upregulating PGC1α, CPT1b, mitochondrial sirtuins (SIRT3-5), and the mitochondrial anti-oxidative genes SOD2 and catalase. In endothelial cells, butyrate suppressed gene expression and LPS-induced secretion of several pro-inflammatory genes, while R-β-hydroxybutyrate acted as a slightly pro-inflammatory molecule. Our observations indicate that butyrate induces transcriptional changes to a higher extent than R-β-hydroxybutyrate in rat myotubes and endothelial cells, in keep with its HDAC inhibitory activity. Also, in contrast with previous reports, R-β-hydroxybutyrate, while inducing histone β-hydroxybutyrylation, did not display a readily detectable HDAC inhibitor activity and exerted a slight pro-inflammatory action on endothelial cells.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
