

Liver Derived FGF21 Maintains Core Body Temperature During Acute Cold Exposure
- PMID: 30679672
- PMCID: PMC6345819
- DOI: 10.1038/s41598-018-37198-y
Liver Derived FGF21 Maintains Core Body Temperature During Acute Cold Exposure
Abstract
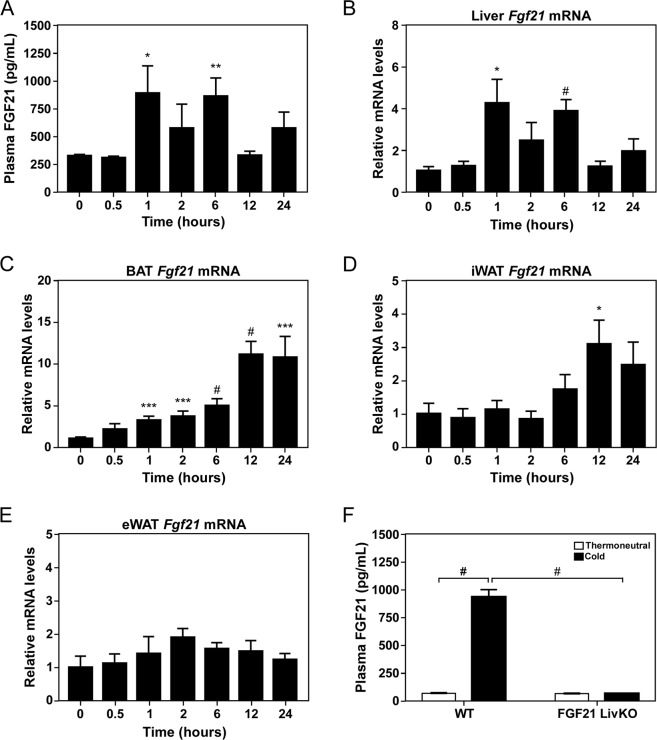
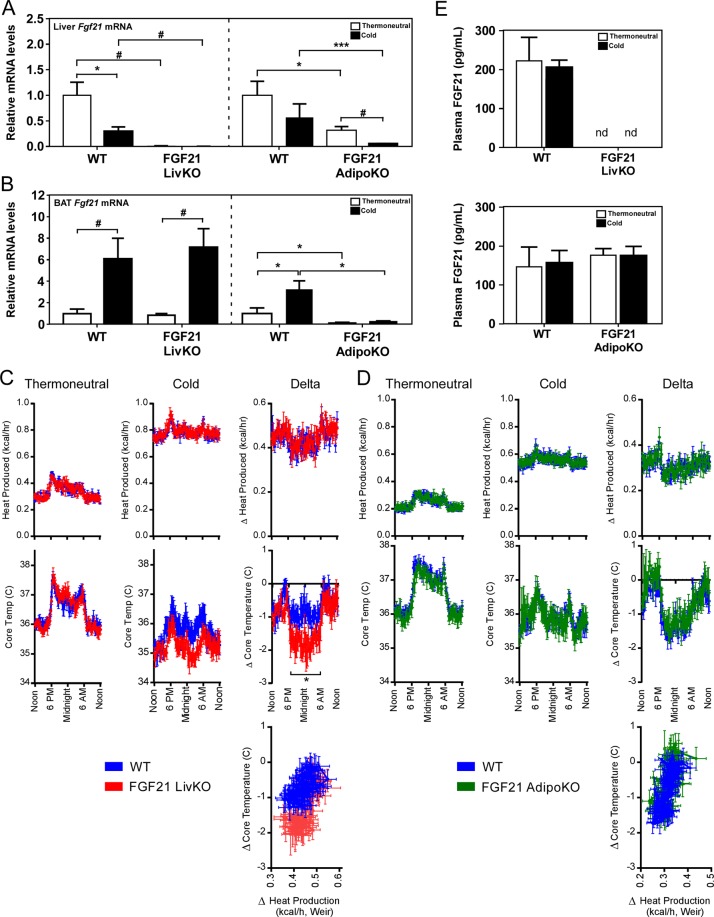
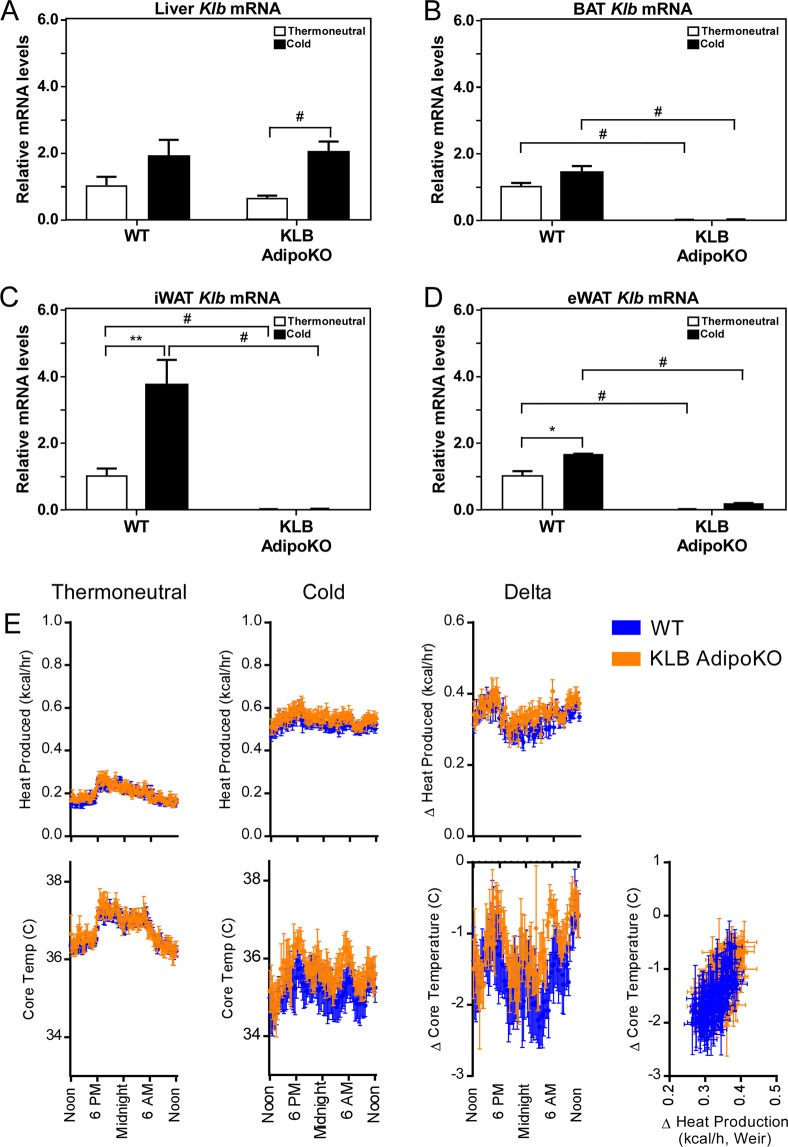
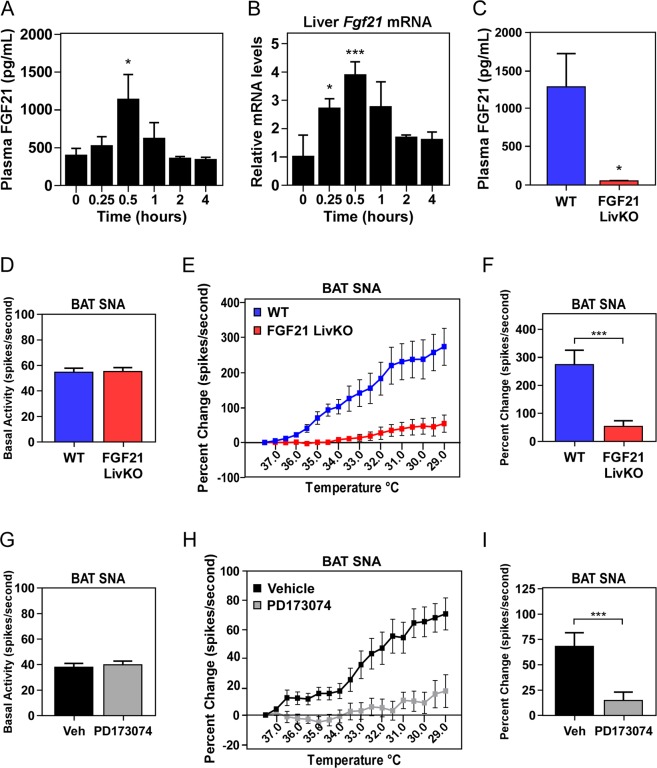
Fibroblast Growth Factor 21 (FGF21) elicits an array of metabolic effects. However, the physiological role of FGF21 during thermal challenges is not clear. In this study, we assessed the tissue source of FGF21 and its site of action to regulate core body temperature in response to cold. Using mice lacking FGF21 specifically in the liver (FGF21 LivKO) or adipose tissues (FGF21 AdipoKO), we performed a series of cold exposure studies to examine the tissue specific induction of FGF21 in response to cold. We also examined the physiological site of FGF21 action during cold exposure by impairing FGF21 signaling to adipose tissues or the central nervous system (CNS) using genetic ablation of the FGF21 co-receptor β-klotho in adipose tissues (KLB AdipoKO) or pharmacological blockage of FGF21 signaling. We found that only liver-derived FGF21 enters circulation during acute cold exposure and is critical for thermoregulation. While FGF21 signaling directly to adipose tissues during cold is dispensable for thermoregulation, central FGF21 signaling is necessary for maximal sympathetic drive to brown adipose tissue to maintain thermoregulation during cold. These data demonstrate a previously unrecognized role for FGF21 in the maintenance of body temperature in response to cold.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
