

Engineering of Chimeric Polyketide Synthases Using SYNZIP Docking Domains
- PMID: 30682239
- PMCID: PMC6554747
- DOI: 10.1021/acschembio.8b01060
Engineering of Chimeric Polyketide Synthases Using SYNZIP Docking Domains
Abstract
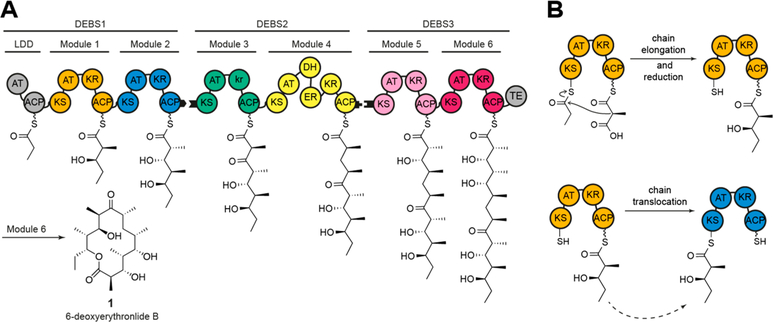
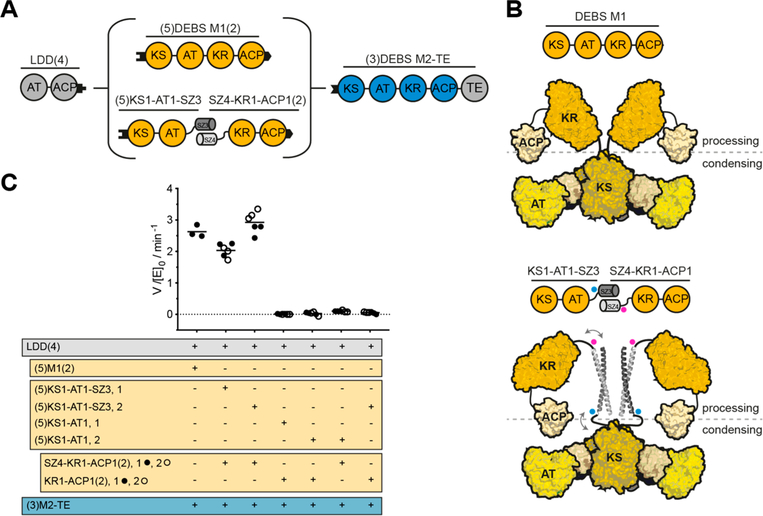
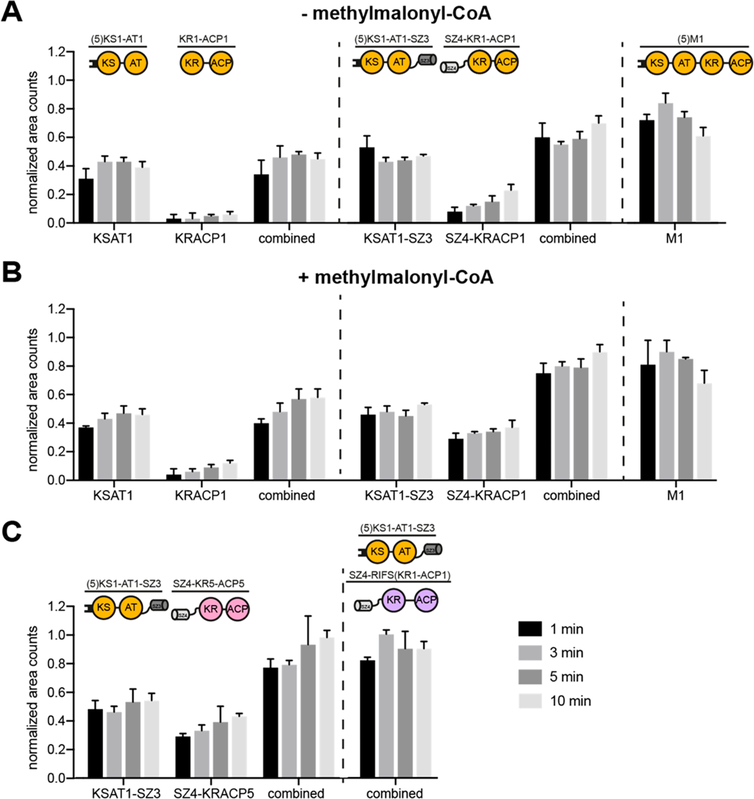
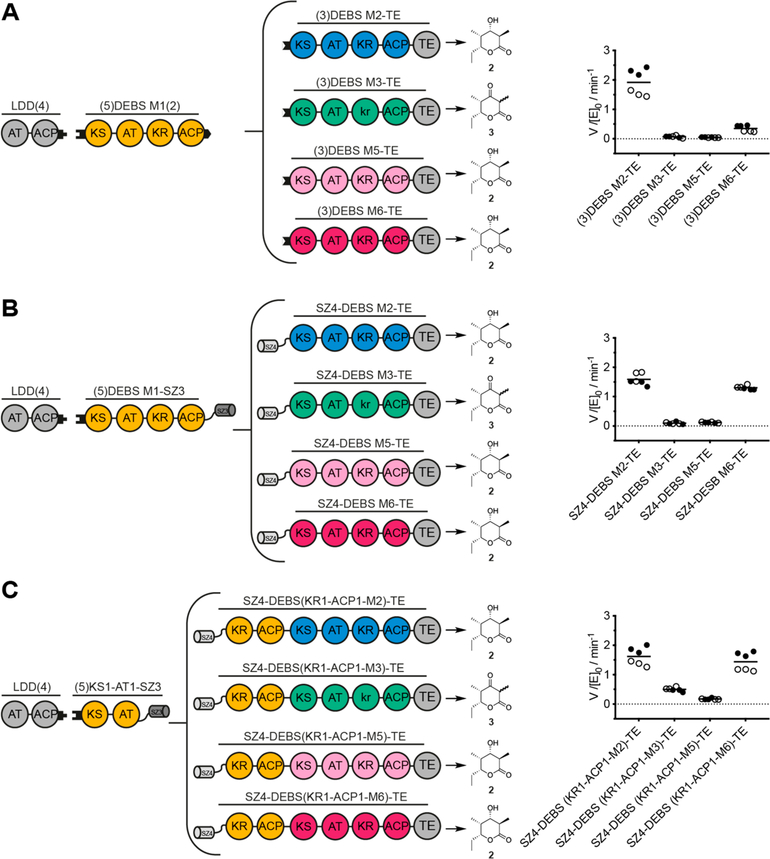
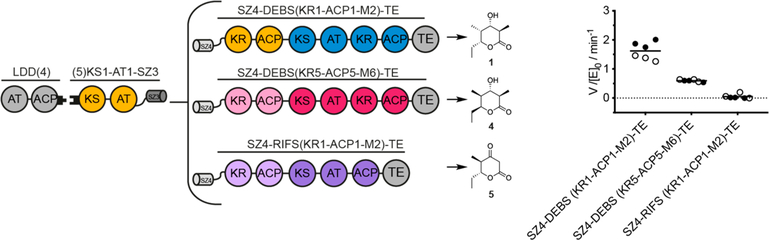
Engineering of assembly line polyketide synthases (PKSs) to produce novel bioactive compounds has been a goal for over 20 years. The apparent modularity of PKSs has inspired many engineering attempts in which entire modules or single domains were exchanged. In recent years, it has become evident that certain domain-domain interactions are evolutionarily optimized and, if disrupted, cause a decrease of the overall turnover rate of the chimeric PKS. In this study, we compared different types of chimeric PKSs in order to define the least invasive interface and to expand the toolbox for PKS engineering. We generated bimodular chimeric PKSs in which entire modules were exchanged, while either retaining a covalent linker between heterologous modules or introducing a noncovalent docking domain, or SYNZIP domain, mediated interface. These chimeric systems exhibited non-native domain-domain interactions during intermodular polyketide chain translocation. They were compared to otherwise equivalent bimodular PKSs in which a noncovalent interface was introduced between the condensing and processing parts of a module, resulting in non-native domain interactions during the extender unit acylation and polyketide chain elongation steps of their catalytic cycles. We show that the natural PKS docking domains can be efficiently substituted with SYNZIP domains and that the newly introduced noncovalent interface between the condensing and processing parts of a module can be harnessed for PKS engineering. Additionally, we established SYNZIP domains as a new tool for engineering PKSs by efficiently bridging non-native interfaces without perturbing PKS activity.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Eng CH, Yuzawa S, Wang G, Baidoo EEK, Katz L, and Keasling JD (2016) Alteration of Polyketide Stereochemistry from anti to syn by a Ketoreductase Domain Exchange in a Type i Modular Polyketide Synthase Subunit. Biochemistry 55, 1677–1680. - PubMed
-
- Yuzawa S, Deng K, Wang G, Baidoo EEK, Northen TR, Adams PD, Katz L, and Keasling JD (2017) Comprehensive in Vitro Analysis of Acyltransferase Domain Exchanges in Modular Polyketide Synthases and Its Application for Short-Chain Ketone Production. ACS Synth. Biol 6, 139–147. - PubMed
-
- Menzella HG, Reid R, Carney JR, Chandran SS, Reisinger SJ, Patel KG, Hopwood DA, and Santi DV (2005) Combinatorial polyketide biosynthesis by de novo design and rearrangement of modular polyketide synthase genes. Nat. Biotechnol 23, 1171–1176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
