

Presynaptic α2δ-2 Calcium Channel Subunits Regulate Postsynaptic GABAA Receptor Abundance and Axonal Wiring
- PMID: 30683685
- PMCID: PMC6445987
- DOI: 10.1523/JNEUROSCI.2234-18.2019
Presynaptic α2δ-2 Calcium Channel Subunits Regulate Postsynaptic GABAA Receptor Abundance and Axonal Wiring
Abstract
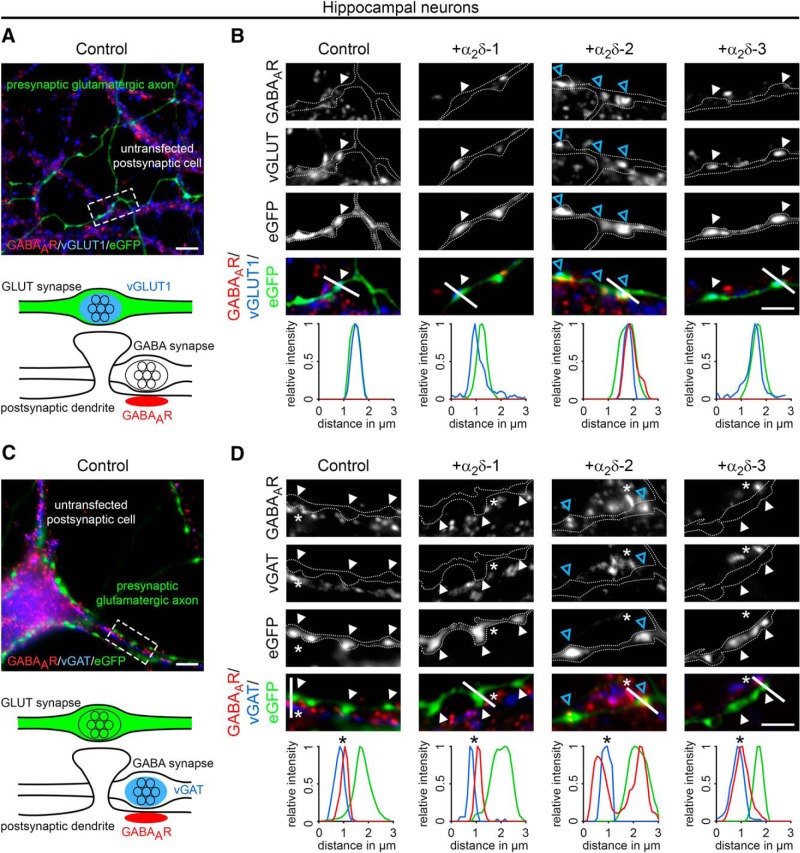
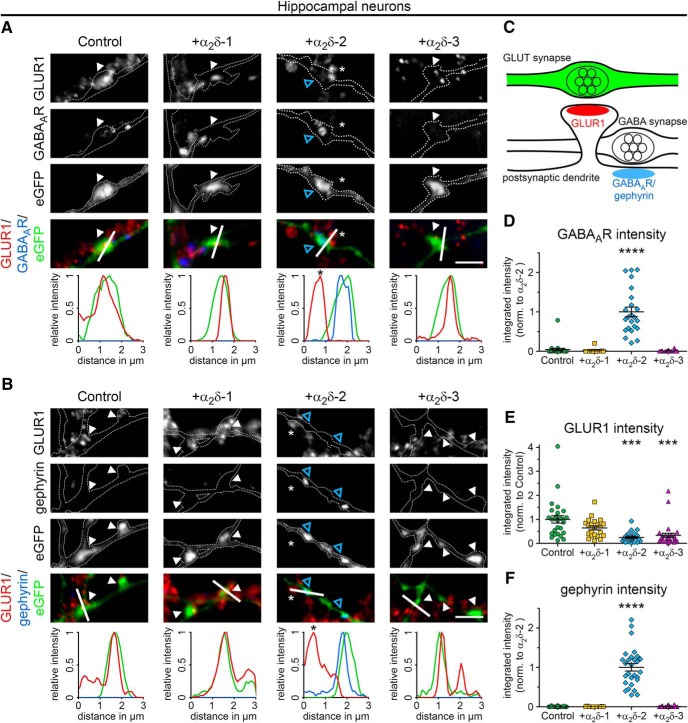
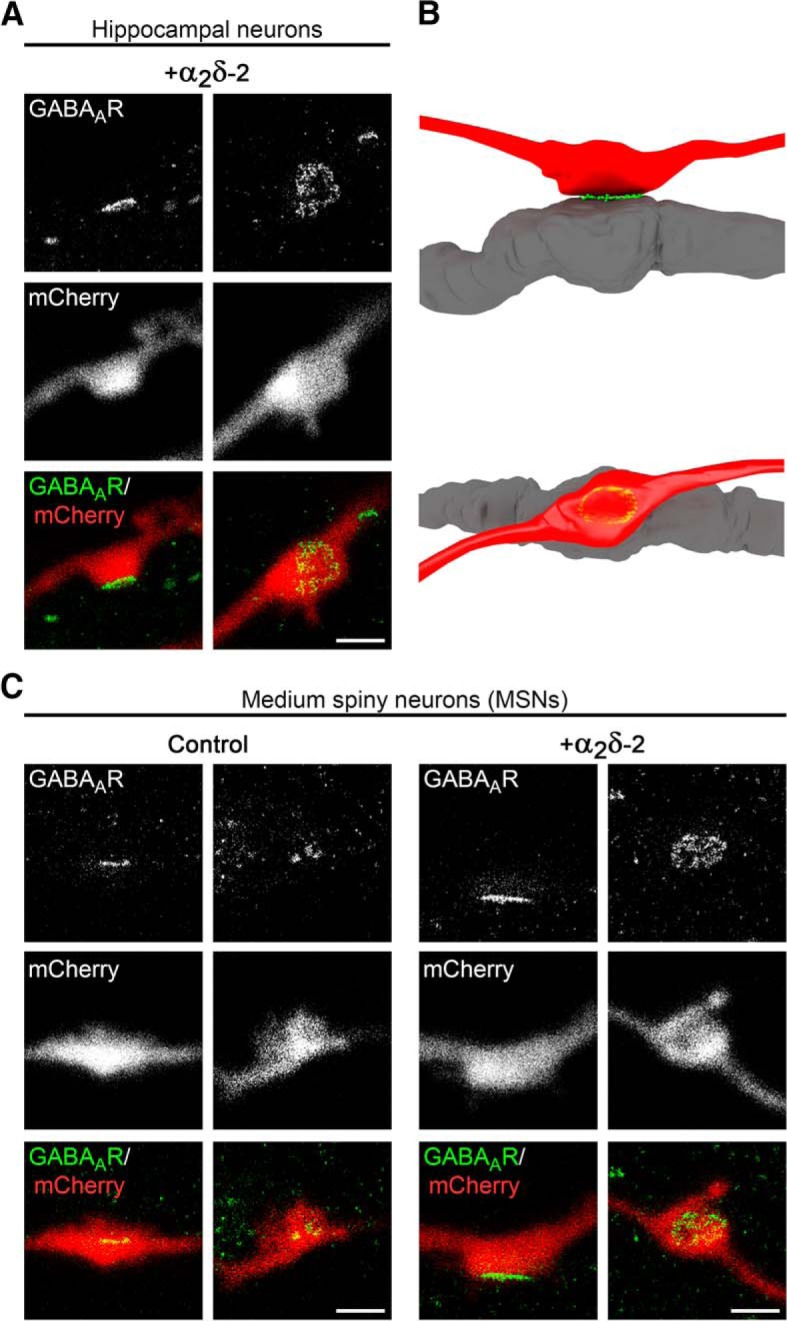
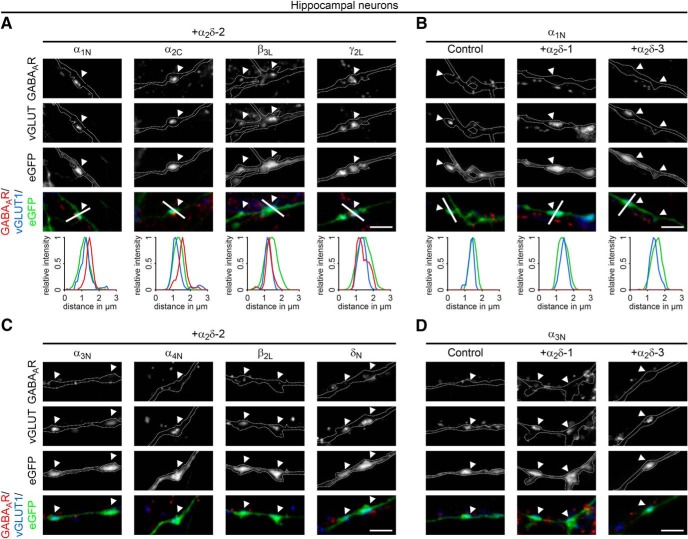
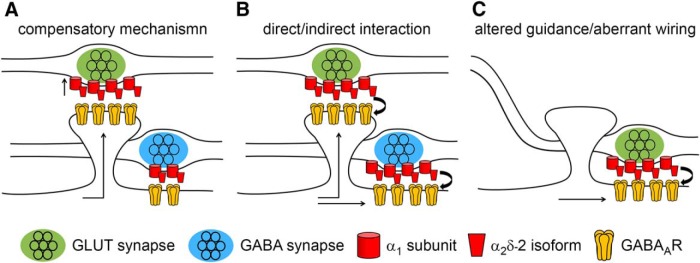
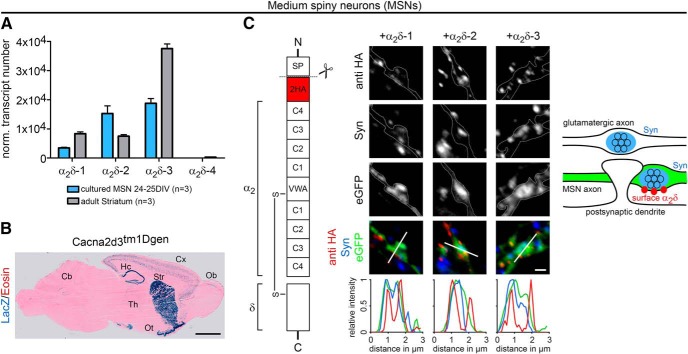
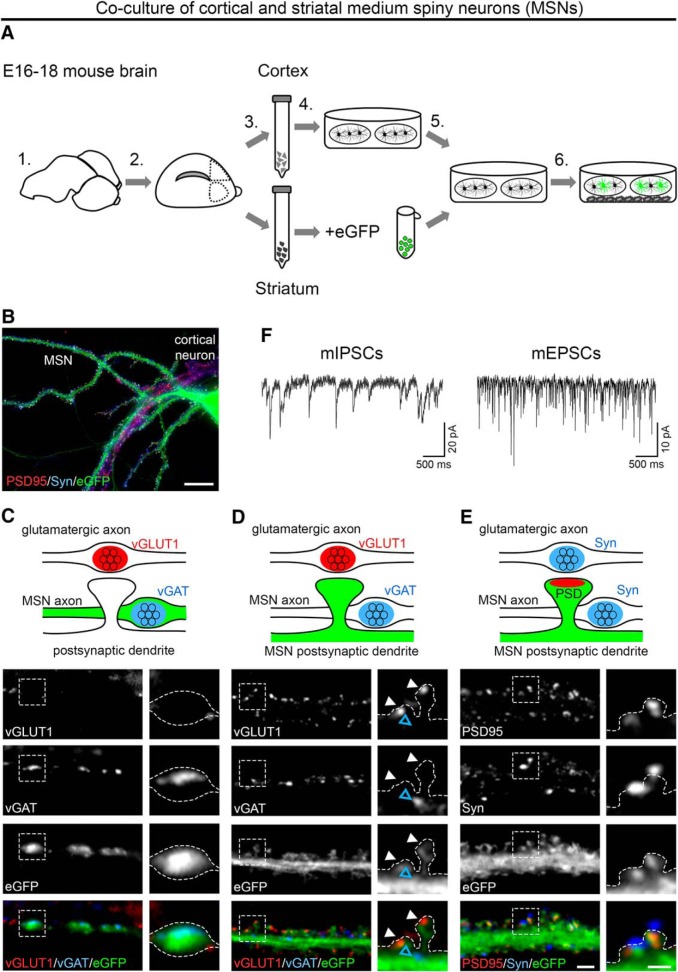
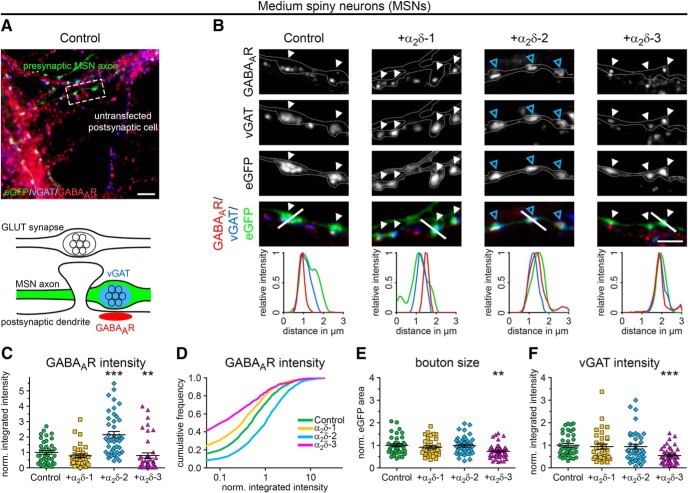
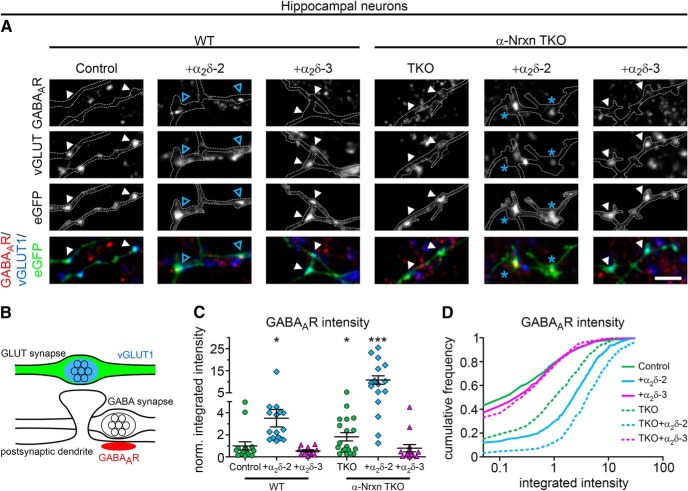
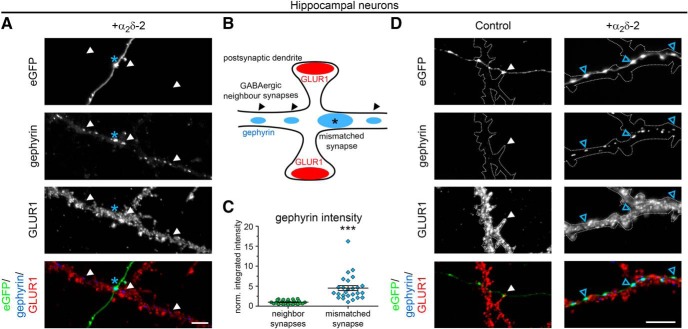
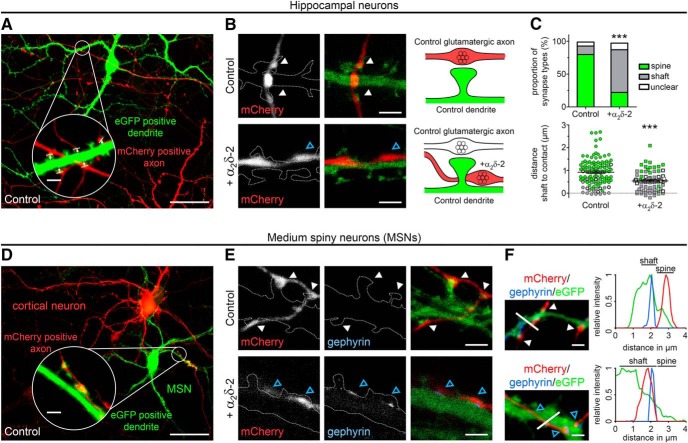
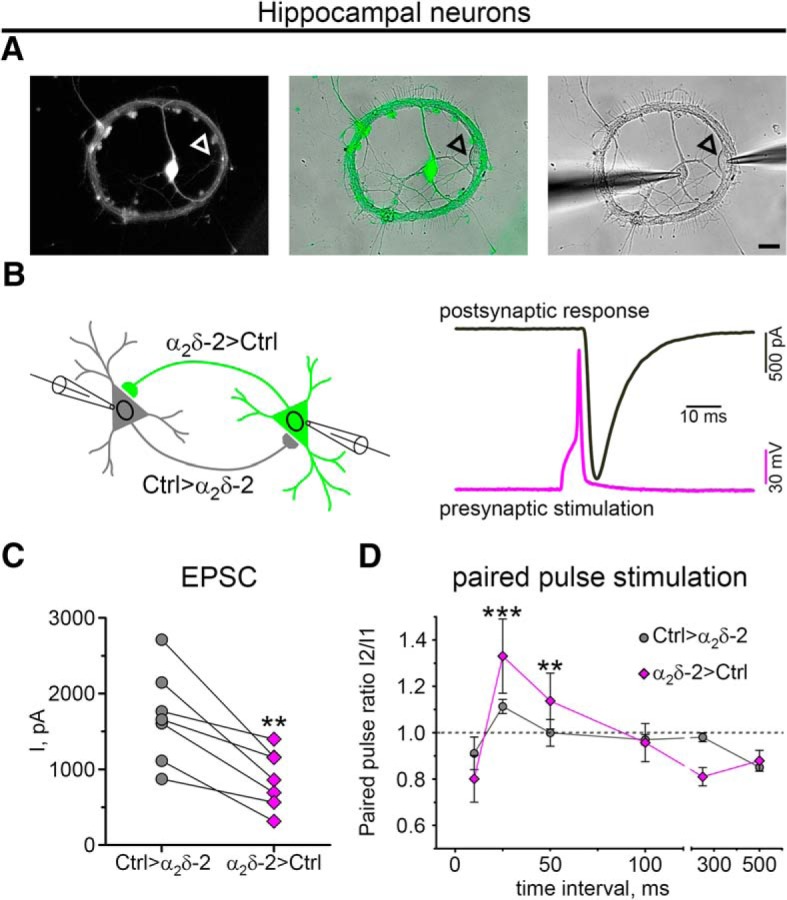
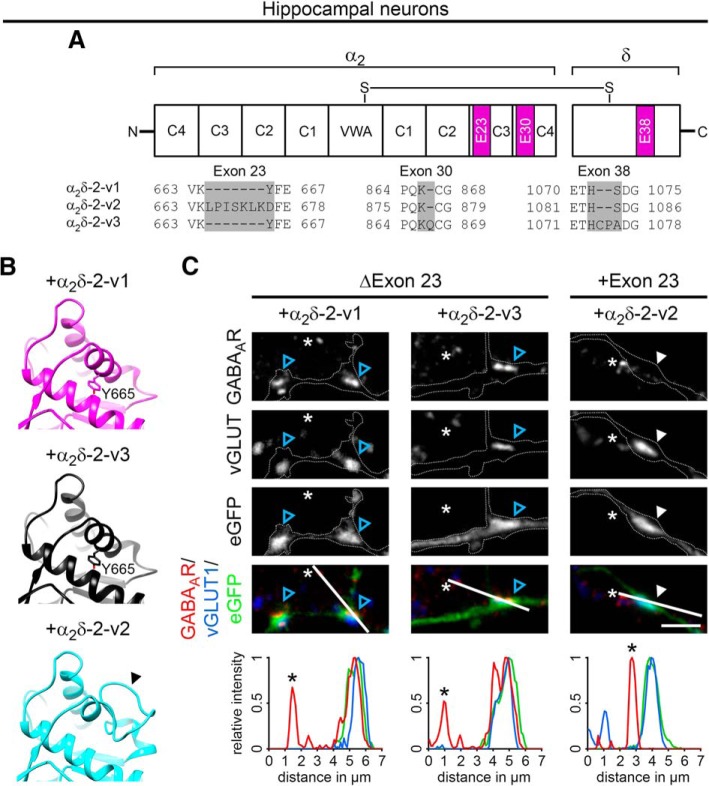
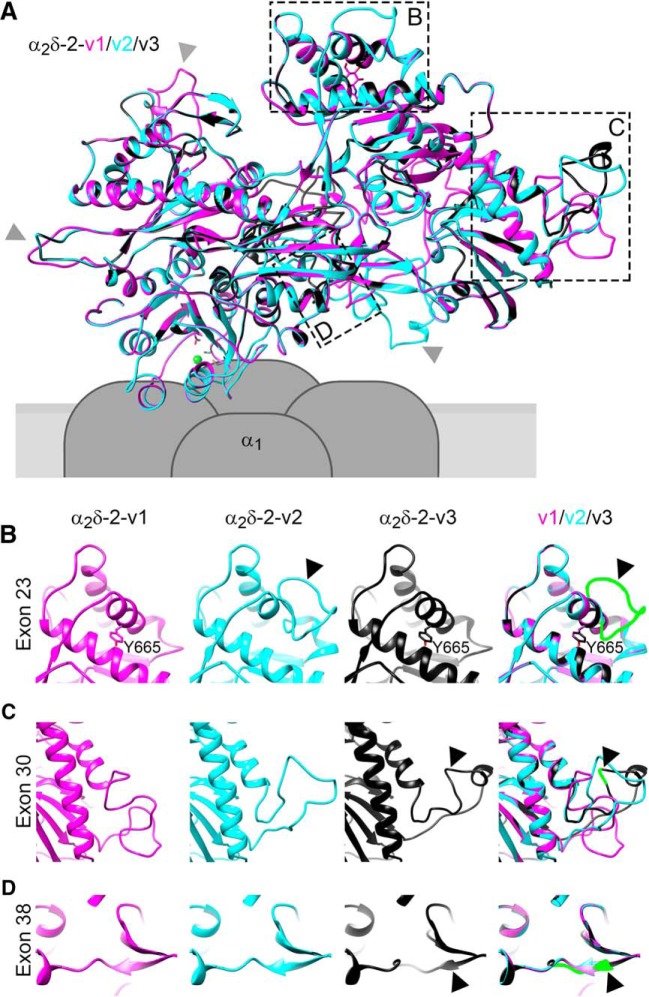
Presynaptic α2δ subunits of voltage-gated calcium channels regulate channel abundance and are involved in glutamatergic synapse formation. However, little is known about the specific functions of the individual α2δ isoforms and their role in GABAergic synapses. Using primary neuronal cultures of embryonic mice of both sexes, we here report that presynaptic overexpression of α2δ-2 in GABAergic synapses strongly increases clustering of postsynaptic GABAARs. Strikingly, presynaptic α2δ-2 exerts the same effect in glutamatergic synapses, leading to a mismatched localization of GABAARs. This mismatching is caused by an aberrant wiring of glutamatergic presynaptic boutons with GABAergic postsynaptic positions. The trans-synaptic effect of α2δ-2 is independent of the prototypical cell-adhesion molecules α-neurexins (α-Nrxns); however, α-Nrxns together with α2δ-2 can modulate postsynaptic GABAAR abundance. Finally, exclusion of the alternatively spliced exon 23 of α2δ-2 is essential for the trans-synaptic mechanism. The novel function of α2δ-2 identified here may explain how abnormal α2δ subunit expression can cause excitatory-inhibitory imbalance often associated with neuropsychiatric disorders.SIGNIFICANCE STATEMENT Voltage-gated calcium channels regulate important neuronal functions such as synaptic transmission. α2δ subunits modulate calcium channels and are emerging as regulators of brain connectivity. However, little is known about how individual α2δ subunits contribute to synapse specificity. Here, we show that presynaptic expression of a single α2δ variant can modulate synaptic connectivity and the localization of inhibitory postsynaptic receptors. Our findings provide basic insights into the development of specific synaptic connections between nerve cells and contribute to our understanding of normal nerve cell functions. Furthermore, the identified mechanism may explain how an altered expression of calcium channel subunits can result in aberrant neuronal wiring often associated with neuropsychiatric disorders such as autism or schizophrenia.
Keywords: auxiliary subunits; cacna2d; cultured hippocampal neurons; imaging; immunocytochemistry; voltage-gated calcium channels.
Copyright © 2019 Geisler et al.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases