

FGL2 promotes tumor progression in the CNS by suppressing CD103+ dendritic cell differentiation
- PMID: 30683885
- PMCID: PMC6347641
- DOI: 10.1038/s41467-018-08271-x
FGL2 promotes tumor progression in the CNS by suppressing CD103+ dendritic cell differentiation
Erratum in
-
Author Correction: FGL2 promotes tumor progression in the CNS by suppressing CD103+ dendritic cell differentiation.Nat Commun. 2019 Feb 15;10(1):862. doi: 10.1038/s41467-019-08770-5. Nat Commun. 2019. PMID: 30770835 Free PMC article.
Abstract
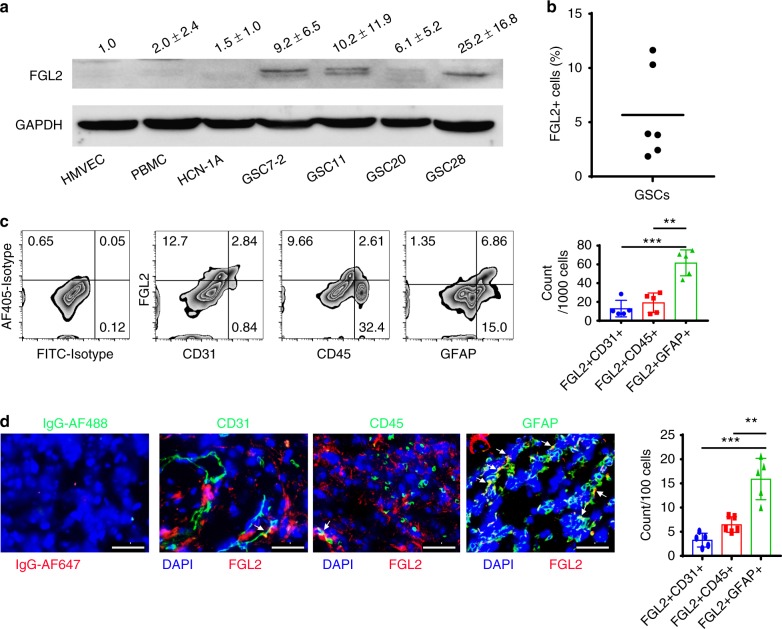
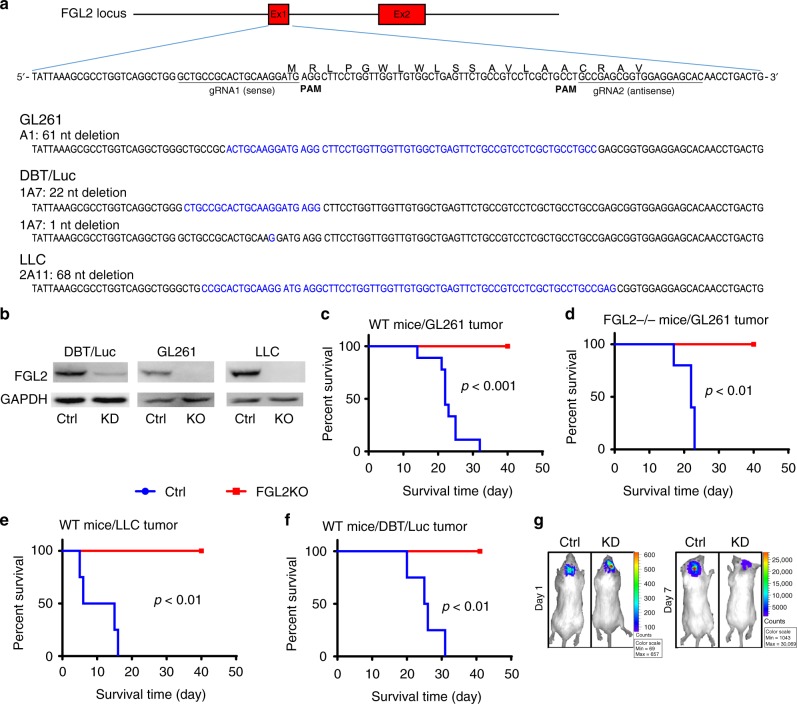
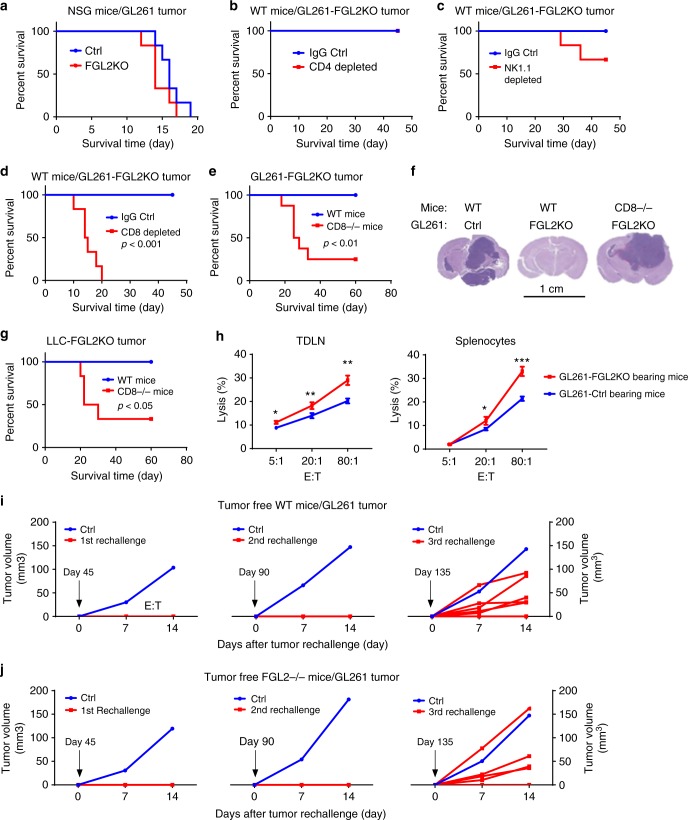
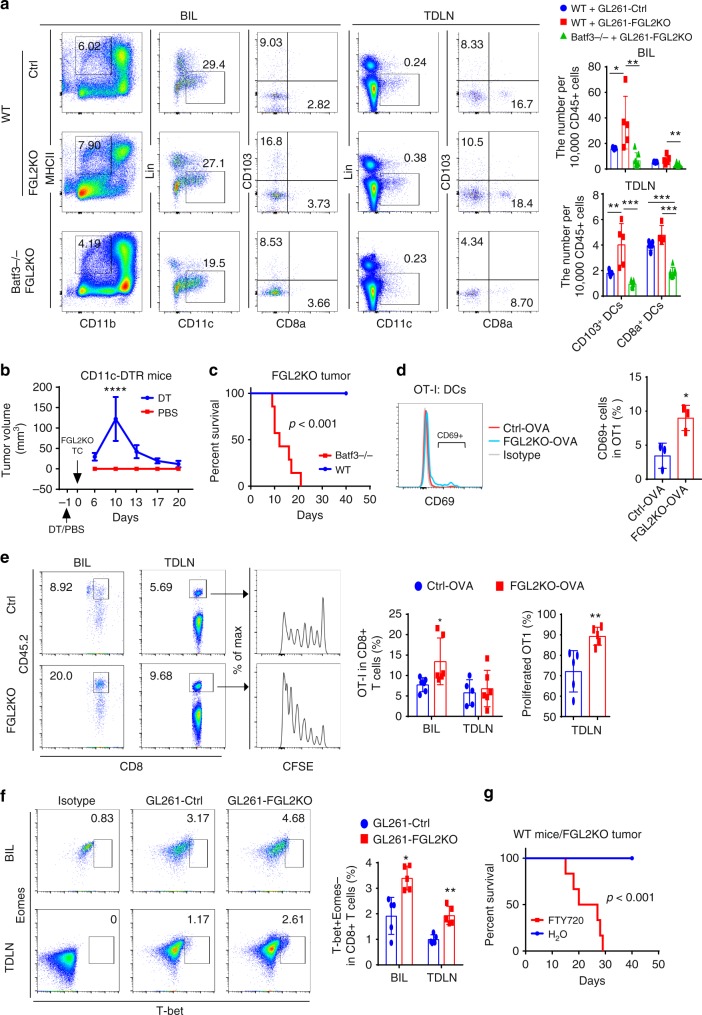
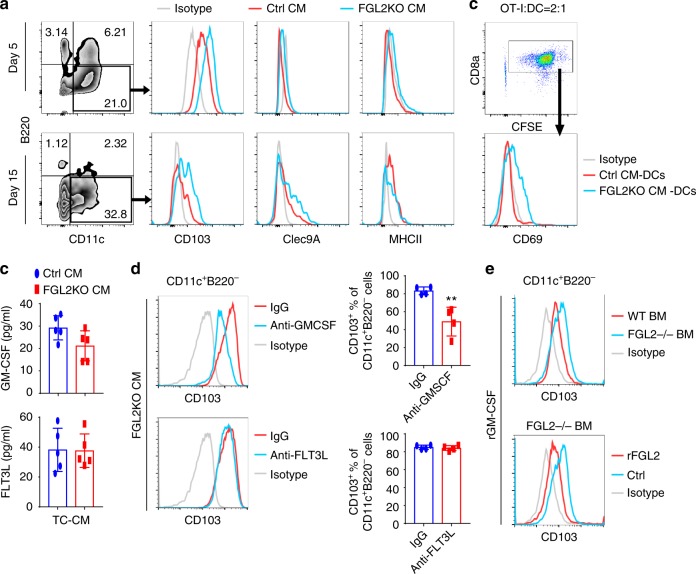
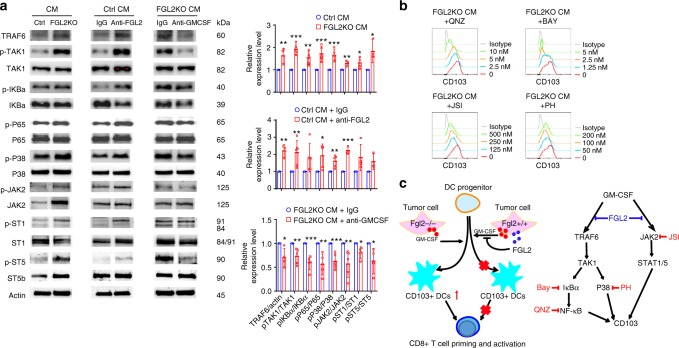
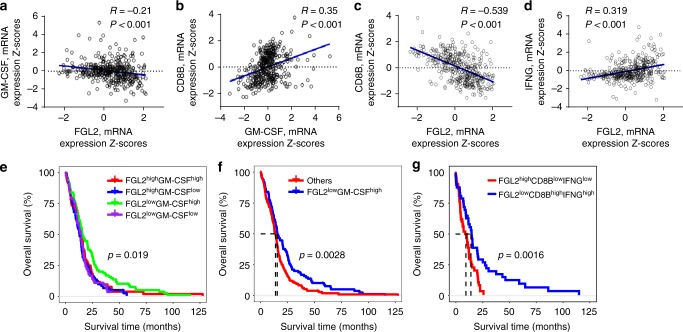
Few studies implicate immunoregulatory gene expression in tumor cells in arbitrating brain tumor progression. Here we show that fibrinogen-like protein 2 (FGL2) is highly expressed in glioma stem cells and primary glioblastoma (GBM) cells. FGL2 knockout in tumor cells did not affect tumor-cell proliferation in vitro or tumor progression in immunodeficient mice but completely impaired GBM progression in immune-competent mice. This impairment was reversed in mice with a defect in dendritic cells (DCs) or CD103+ DC differentiation in the brain and in tumor-draining lymph nodes. The presence of FGL2 in tumor cells inhibited granulocyte-macrophage colony-stimulating factor (GM-CSF)-induced CD103+ DC differentiation by suppressing NF-κB, STAT1/5, and p38 activation. These findings are relevant to GBM patients because a low level of FGL2 expression with concurrent high GM-CSF expression is associated with higher CD8B expression and longer survival. These data provide a rationale for therapeutic inhibition of FGL2 in brain tumors.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
