

Correcting aberrant kinetochore microtubule attachments: a hidden regulation of Aurora B on microtubules
- PMID: 30684807
- PMCID: PMC6646109
- DOI: 10.1016/j.ceb.2018.12.007
Correcting aberrant kinetochore microtubule attachments: a hidden regulation of Aurora B on microtubules
Abstract
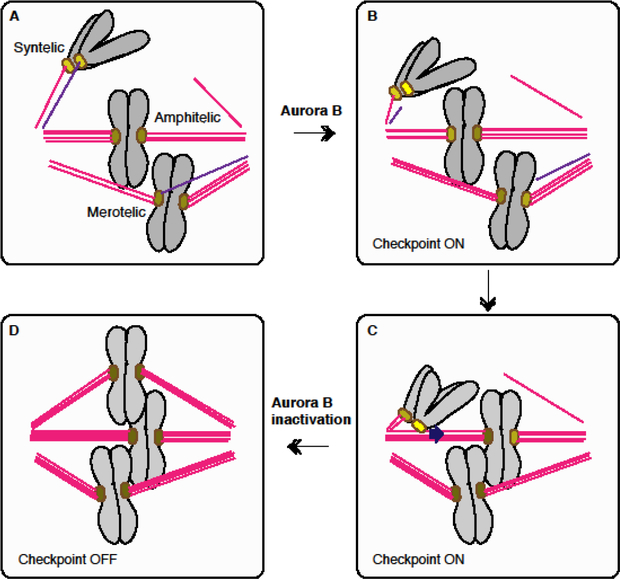
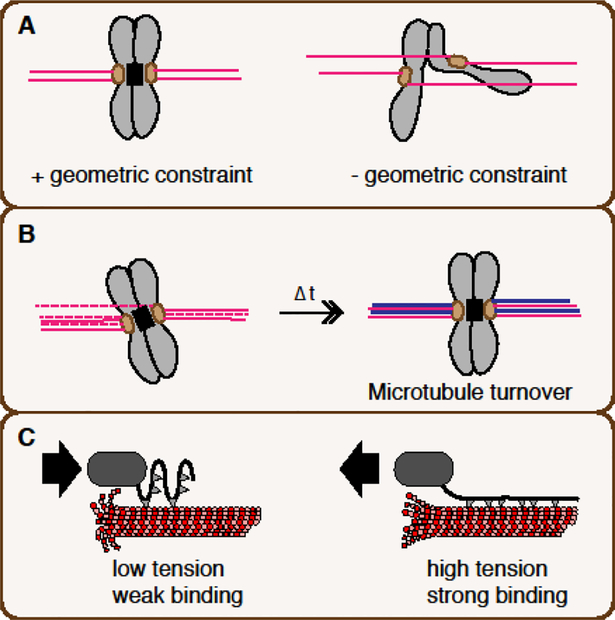
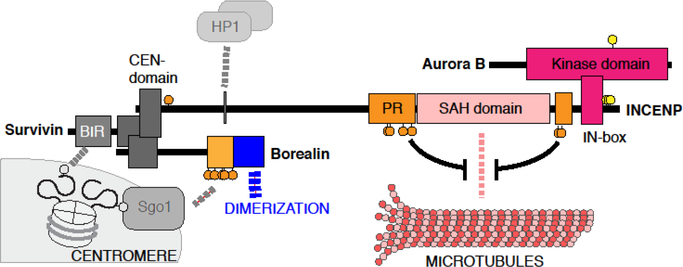
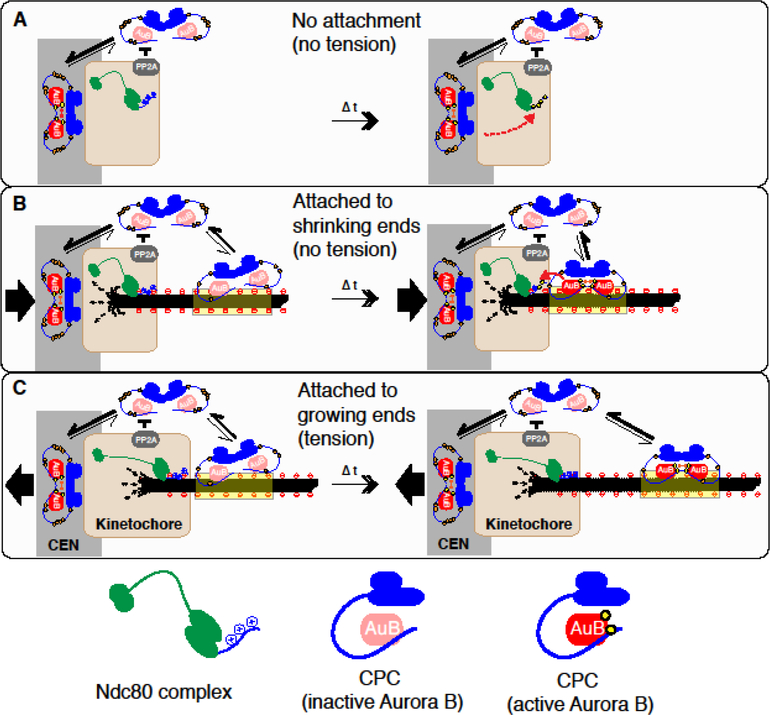
For equal chromosome segregation, a pair of kinetochores on each duplicated chromosome must attach to microtubules connecting to opposite poles. The protein kinase Aurora B plays a critical role in destabilizing microtubules attached in a wrong orientation through phosphorylating kinetochore proteins. The mechanism behind this selective destabilization of aberrant attachments remains elusive. While Aurora B is most enriched on the centromere from prophase to metaphase, emerging evidence suggests the importance of Aurora B on microtubules in this process. Here I discuss two hypothetical models that could explain the requirement of Aurora B on microtubules for selective destabilization of aberrant attachments; microtubule-induced substrate masking and treadmill-removal of Aurora B on microtubules proximal to polymerizing ends.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Aurora A kinase phosphorylates Hec1 to regulate metaphase kinetochore-microtubule dynamics.J Cell Biol. 2018 Jan 2;217(1):163-177. doi: 10.1083/jcb.201707160. Epub 2017 Nov 29. J Cell Biol. 2018. PMID: 29187526 Free PMC article.
-
Regulation of kinetochore-microtubule attachments by Aurora B kinase.Biochem Soc Trans. 2009 Oct;37(Pt 5):976-80. doi: 10.1042/BST0370976. Biochem Soc Trans. 2009. PMID: 19754435 Review.
-
Aurora B/AIR-2 regulates sister centromere resolution and CENP-A/HCP-3 organization to prevent merotelic attachments.J Mol Cell Biol. 2025 May 2;16(10):mjae045. doi: 10.1093/jmcb/mjae045. J Mol Cell Biol. 2025. PMID: 39415429 Free PMC article.
-
Regulatory mechanisms of kinetochore-microtubule interaction in mitosis.Cell Mol Life Sci. 2013 Feb;70(4):559-79. doi: 10.1007/s00018-012-1057-7. Epub 2012 Jul 4. Cell Mol Life Sci. 2013. PMID: 22752158 Free PMC article. Review.
-
A tension-independent mechanism reduces Aurora B-mediated phosphorylation upon microtubule capture by CENP-E at the kinetochore.Cell Cycle. 2019 Jun;18(12):1349-1363. doi: 10.1080/15384101.2019.1617615. Epub 2019 May 23. Cell Cycle. 2019. PMID: 31122175 Free PMC article.
Cited by
-
Centromere drive: model systems and experimental progress.Chromosome Res. 2022 Sep;30(2-3):187-203. doi: 10.1007/s10577-022-09696-3. Epub 2022 Jun 22. Chromosome Res. 2022. PMID: 35731424 Free PMC article. Review.
-
CCAR2 controls mitotic progression through spatiotemporal regulation of Aurora B.Cell Death Dis. 2022 Jun 7;13(6):534. doi: 10.1038/s41419-022-04990-8. Cell Death Dis. 2022. PMID: 35672287 Free PMC article.
-
Shake It Off: The Elimination of Erroneous Kinetochore-Microtubule Attachments and Chromosome Oscillation.Int J Mol Sci. 2021 Mar 20;22(6):3174. doi: 10.3390/ijms22063174. Int J Mol Sci. 2021. PMID: 33804687 Free PMC article. Review.
-
Epigenetic rewriting at centromeric DNA repeats leads to increased chromatin accessibility and chromosomal instability.Epigenetics Chromatin. 2021 Jul 28;14(1):35. doi: 10.1186/s13072-021-00410-x. Epigenetics Chromatin. 2021. PMID: 34321103 Free PMC article.
-
Borealin directs recruitment of the CPC to oocyte chromosomes and movement to the microtubules.J Cell Biol. 2021 Jun 7;220(6):e202006018. doi: 10.1083/jcb.202006018. J Cell Biol. 2021. PMID: 33836043 Free PMC article.
References
-
- Kitajima TS, Ohsugi M, Ellenberg J: Complete kinetochore tracking reveals error-prone homologous chromosome biorientation in mammalian oocytes. Cell 2011, 146:568–581. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
