

Loss of SLC25A11 causes suppression of NSCLC and melanoma tumor formation
- PMID: 30686754
- PMCID: PMC6413681
- DOI: 10.1016/j.ebiom.2019.01.036
Loss of SLC25A11 causes suppression of NSCLC and melanoma tumor formation
Abstract
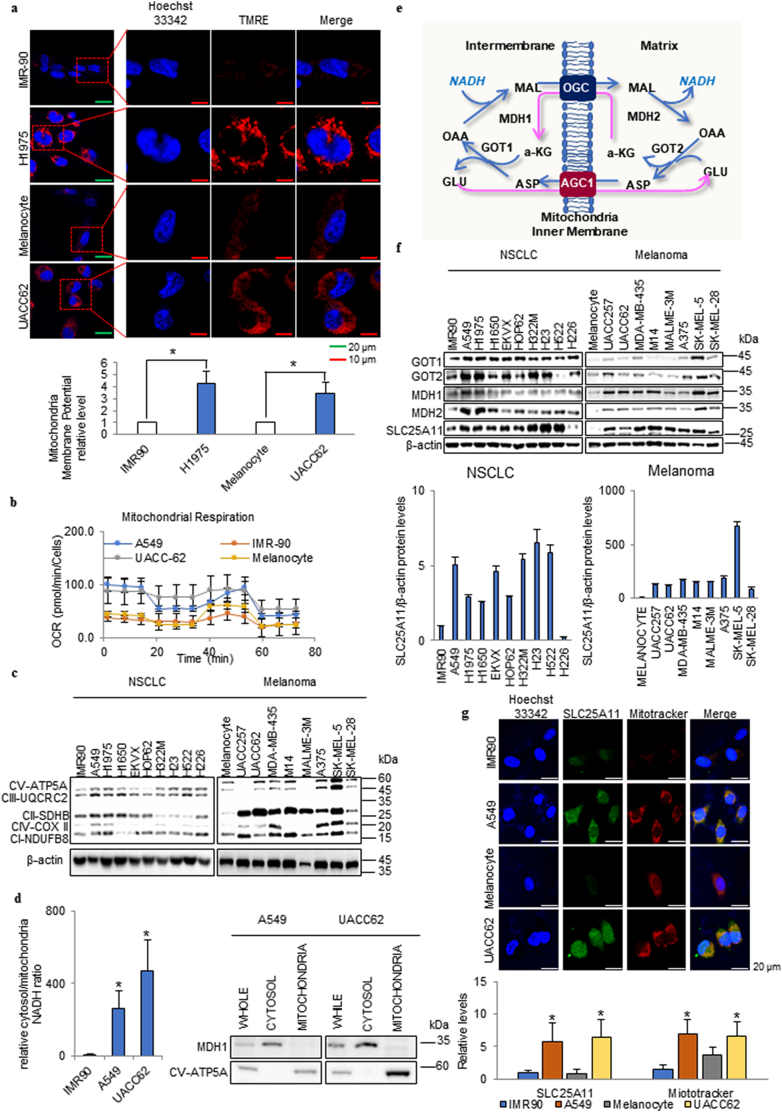
Background: Fast growing cancer cells require greater amounts of ATP than normal cells. Although glycolysis was suggested as a source of anabolic metabolism based on lactate production, the main source of ATP to support cancer cell metabolism remains unidentified.
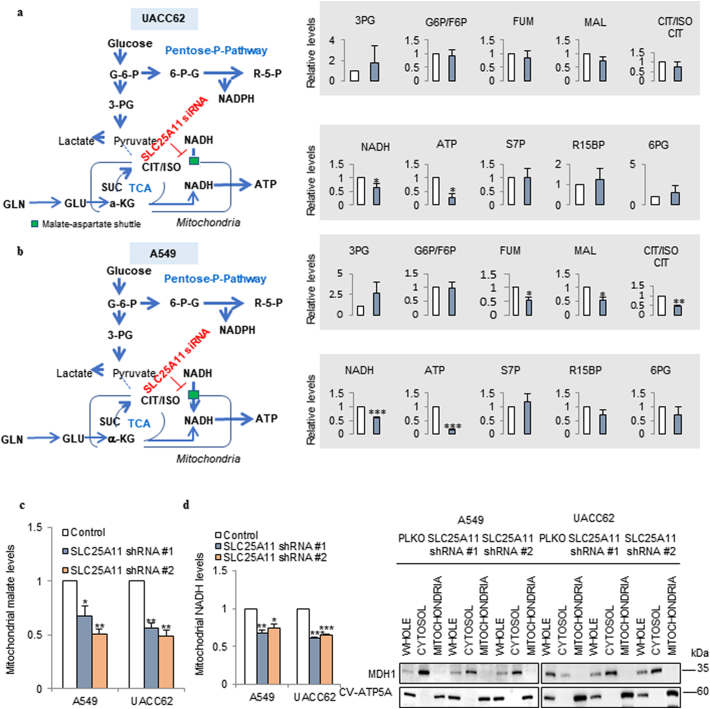
Methods: We have proposed that the oxoglutarate carrier SLC25A11 is important for ATP production in cancer by NADH transportation from the cytosol to mitochondria as a malate. We have examined not only changes of ATP and NADH but also changes of metabolites after SLC25A11 knock down in cancer cells.
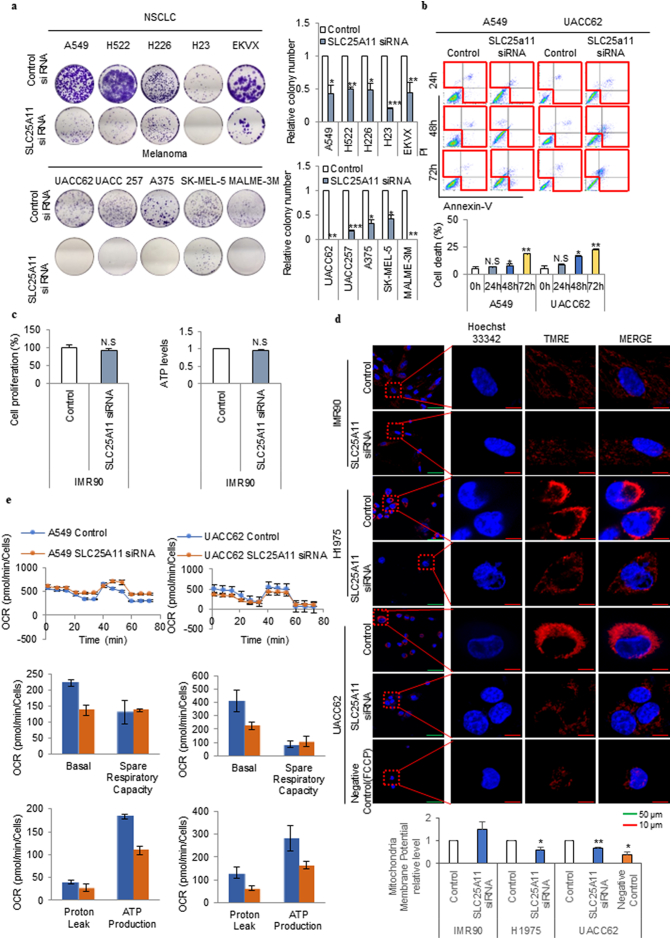
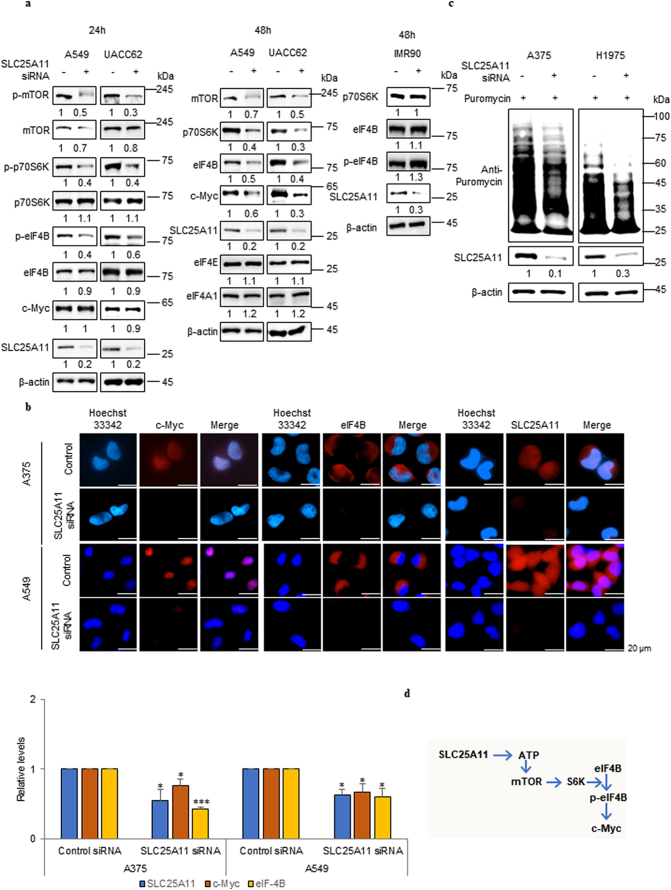
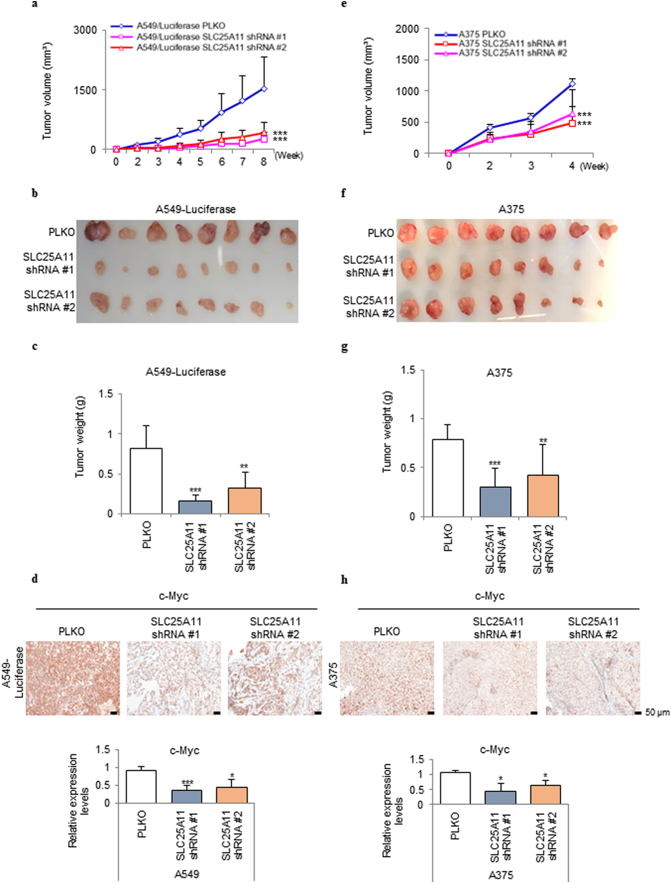
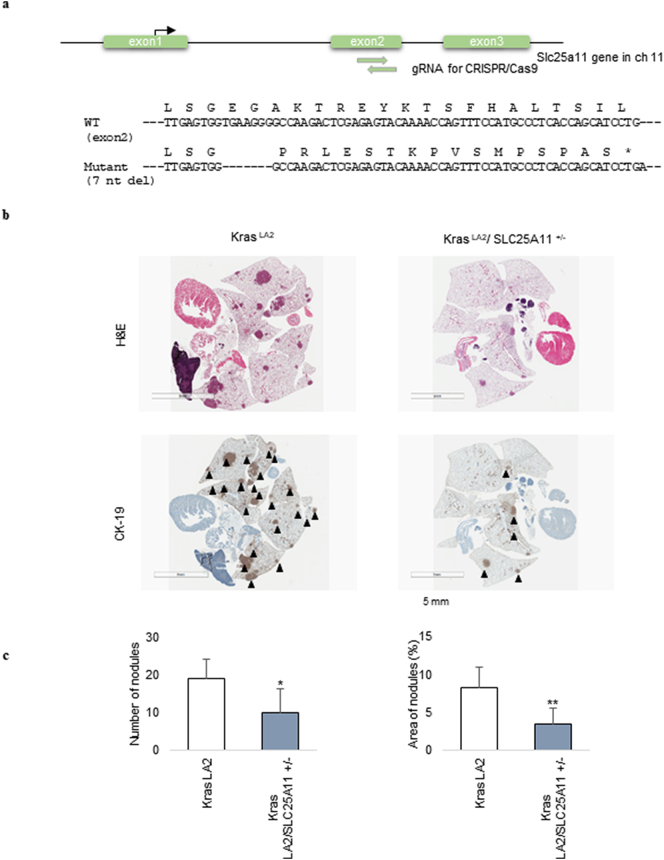
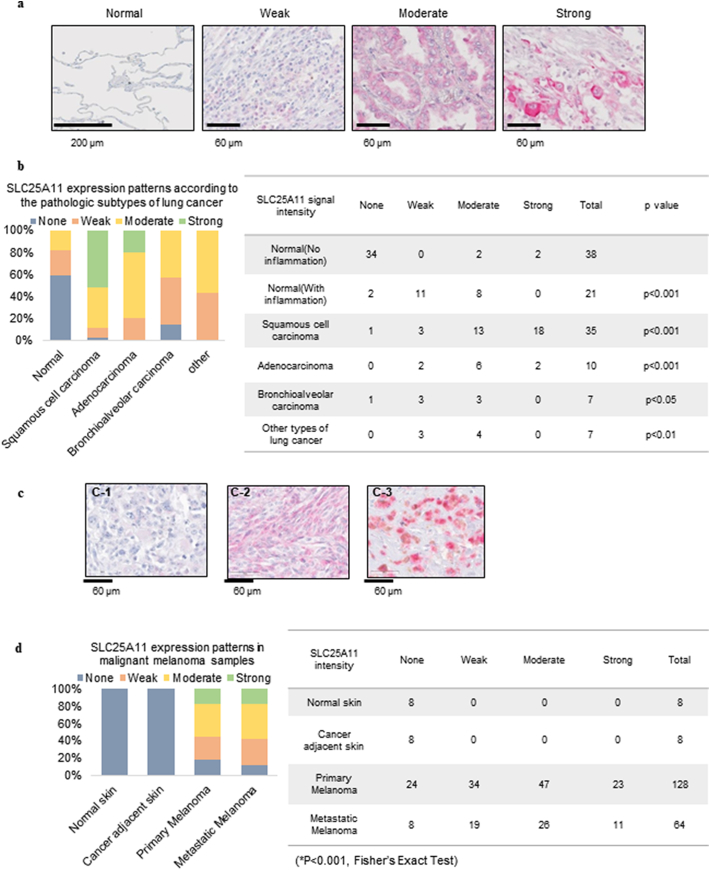
Findings: The mitochondrial electron transport chain was functionally active in cancer cells. The cytosolic to mitochondrial NADH ratio was higher in non-small cell lung cancer (NSCLC) and melanoma cells than in normal cells. This was consistent with higher levels of the oxoglutarate carrier SLC25A11. Blocking malate transport by knockdown of SLC25A11 significantly impaired ATP production and inhibited the growth of cancer cells, which was not observed in normal cells. In in vivo experiments, heterozygote of SLC25A11 knock out mice suppressed KRASLA2 lung tumor formation by cross breeding.
Interpretation: Cancer cells critically depended on the oxoglutarate carrier SLC25A11 for transporting NADH from cytosol to mitochondria as a malate form for the purpose of ATP production. Therefore blocking SLC25A11 may have an advantage in stopping cancer growth by reducing ATP production. FUND: The Basic Science Research Program through the National Research Foundation of Korea funded by the Ministry of Science and ICT to SYK (NRF-2017R1A2B2003428).
Keywords: Cancer metabolism; Cancer therapeutic target; Malate aspartate shuttle; Oxoglutarate carrier; SLC25A11.
Copyright © 2019 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Warburg O. On respiratory impairment in cancer cells. Science. 1956;124(3215):269–270. - PubMed
-
- Weinhouse S. On respiratory impairment in cancer cells. Science. 1956;124(3215):267–269. - PubMed
-
- Smolkova K., Bellance N., Scandurra F., Genot E., Gnaiger E., Plecita-Hlavata L. Mitochondrial bioenergetic adaptations of breast cancer cells to aglycemia and hypoxia. J Bioenerg Biomembr. 2010;42(1):55–67. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
