

Cholesterol Metabolism Is a Druggable Axis that Independently Regulates Tau and Amyloid-β in iPSC-Derived Alzheimer's Disease Neurons
- PMID: 30686764
- PMCID: PMC6414424
- DOI: 10.1016/j.stem.2018.12.013
Cholesterol Metabolism Is a Druggable Axis that Independently Regulates Tau and Amyloid-β in iPSC-Derived Alzheimer's Disease Neurons
Abstract
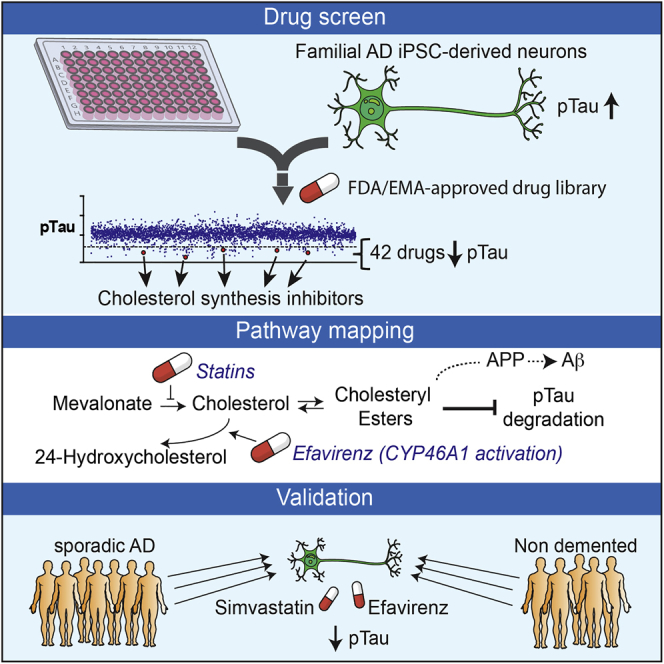
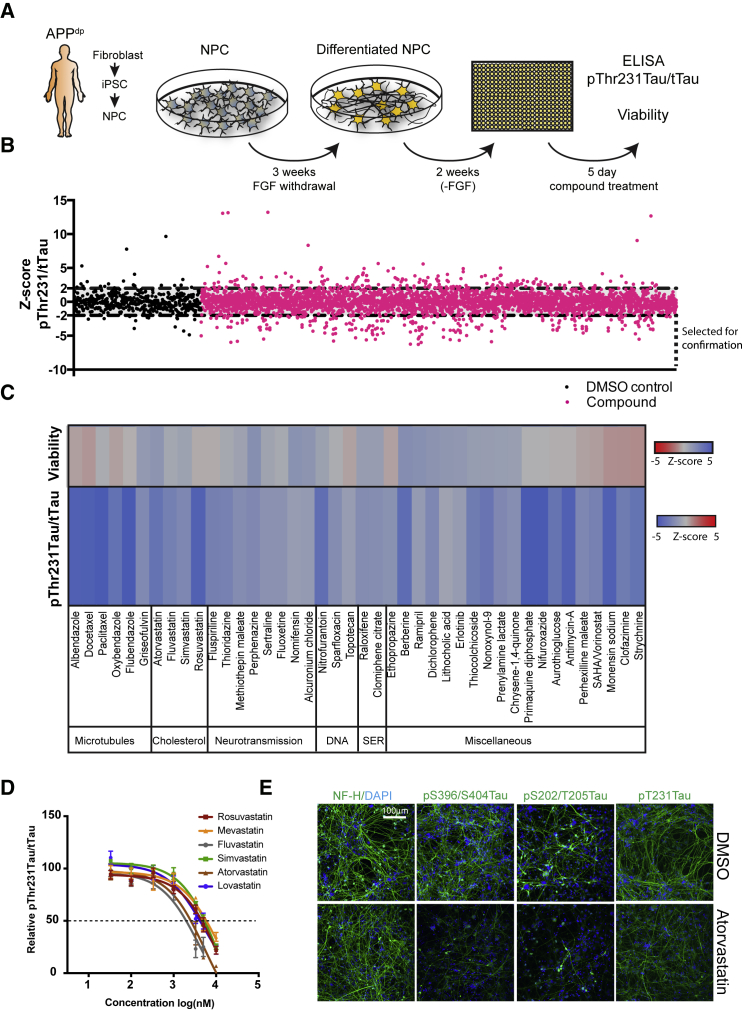
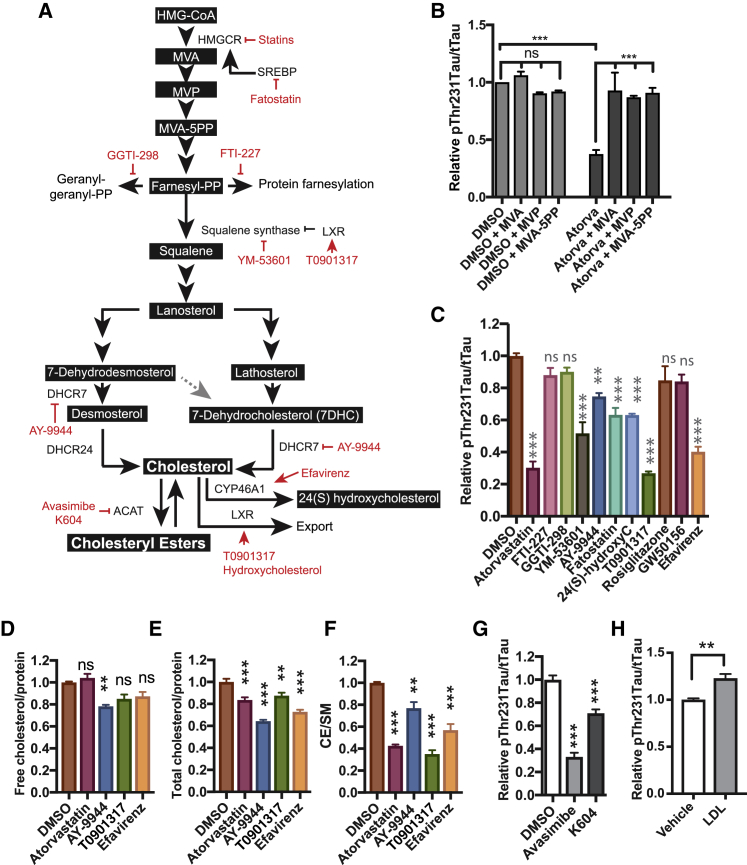
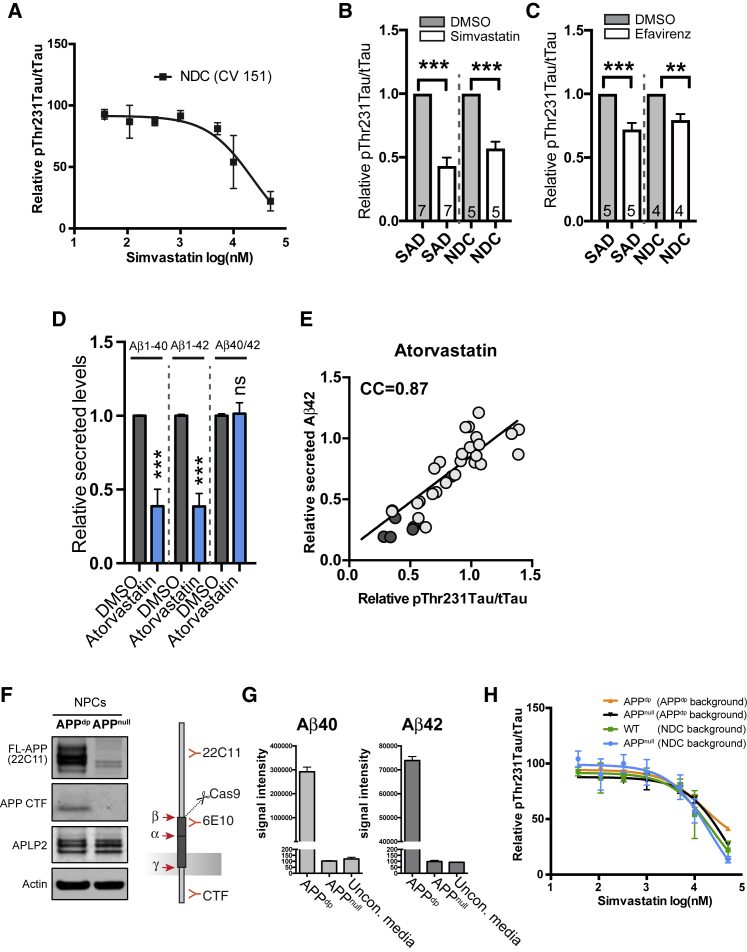
Genetic, epidemiologic, and biochemical evidence suggests that predisposition to Alzheimer's disease (AD) may arise from altered cholesterol metabolism, although the molecular pathways that may link cholesterol to AD phenotypes are only partially understood. Here, we perform a phenotypic screen for pTau accumulation in AD-patient iPSC-derived neurons and identify cholesteryl esters (CE), the storage product of excess cholesterol, as upstream regulators of Tau early during AD development. Using isogenic induced pluripotent stem cell (iPSC) lines carrying mutations in the cholesterol-binding domain of APP or APP null alleles, we found that while CE also regulate Aβ secretion, the effects of CE on Tau and Aβ are mediated by independent pathways. Efficacy and toxicity screening in iPSC-derived astrocytes and neurons showed that allosteric activation of CYP46A1 lowers CE specifically in neurons and is well tolerated by astrocytes. These data reveal that CE independently regulate Tau and Aβ and identify a druggable CYP46A1-CE-Tau axis in AD.
Keywords: Alzheimer’s disease; CYP46A1 Tau; amyloid beta; cholesterol metabolism; cholesteryl esters; disease modeling; drug screening; induced pluripotent stem cells; lipids; proteostasis.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
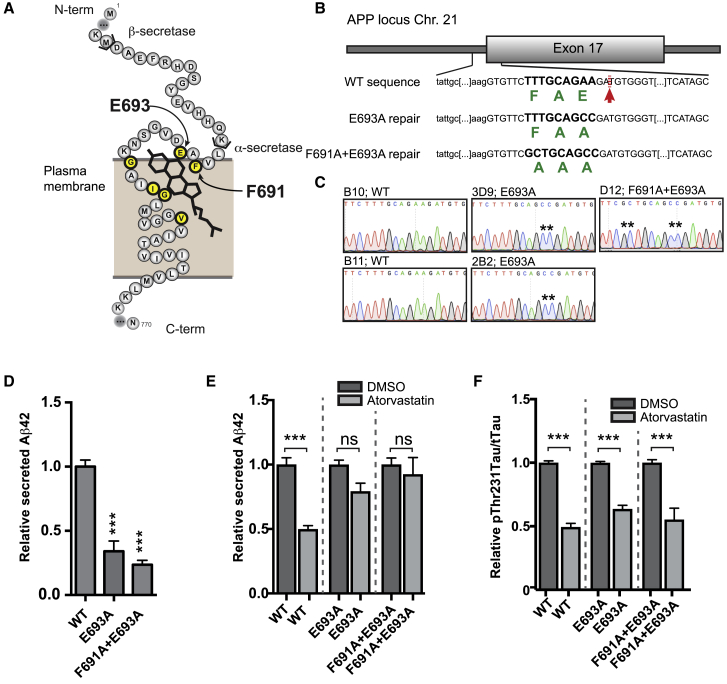
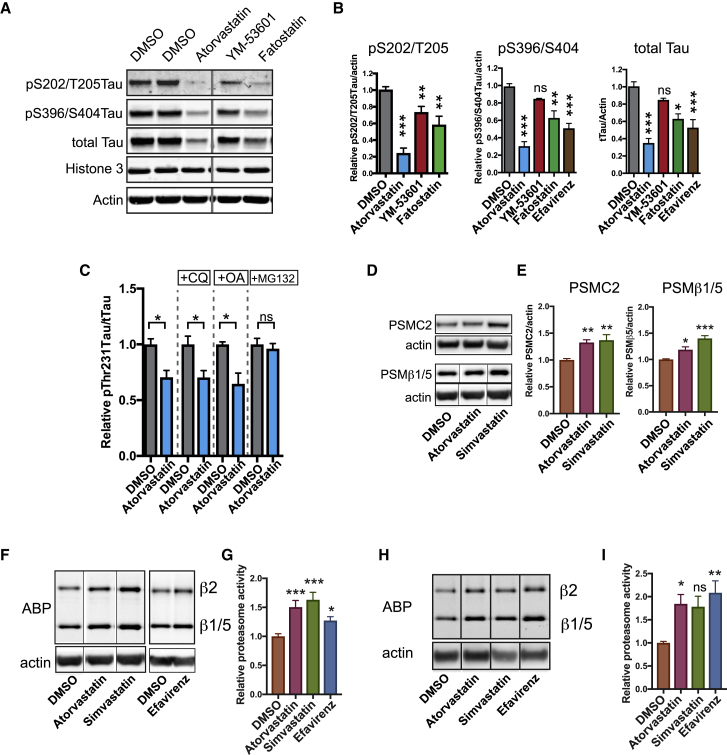
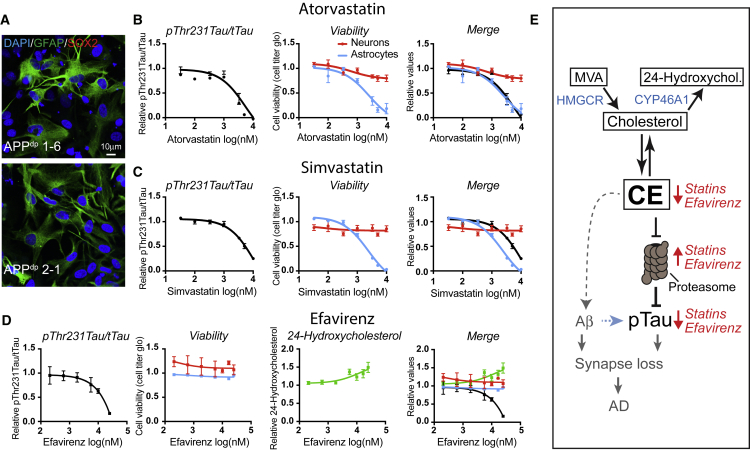
Comment in
-
Unraveling the Paradox of Statins with Human Neurons: New Leads in Alzheimer's Disease.Cell Stem Cell. 2019 Mar 7;24(3):347-349. doi: 10.1016/j.stem.2019.02.003. Cell Stem Cell. 2019. PMID: 30849361
References
-
- Apostolova N., Blas-Garcia A., Galindo M.J., Esplugues J.V. Efavirenz: what is known about the cellular mechanisms responsible for its adverse effects. Eur. J. Pharmacol. 2017;812:163–173. - PubMed
-
- Berkers C.R., van Leeuwen F.W.B., Groothuis T.A., Peperzak V., van Tilburg E.W., Borst J., Neefjes J.J., Ovaa H. Profiling proteasome activity in tissue with fluorescent probes. Mol. Pharm. 2007;4:739–748. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
