

Characterization of Non-hormone Expressing Endocrine Cells in Fetal and Infant Human Pancreas
- PMID: 30687234
- PMCID: PMC6334491
- DOI: 10.3389/fendo.2018.00791
Characterization of Non-hormone Expressing Endocrine Cells in Fetal and Infant Human Pancreas
Abstract
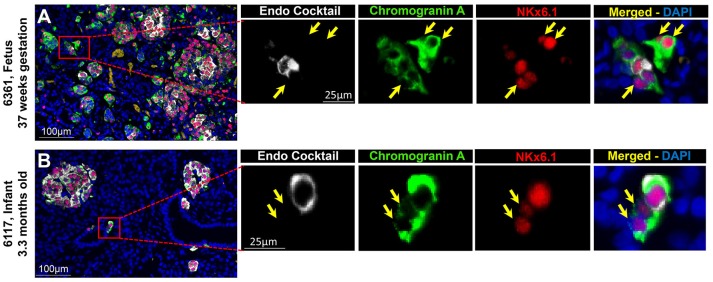
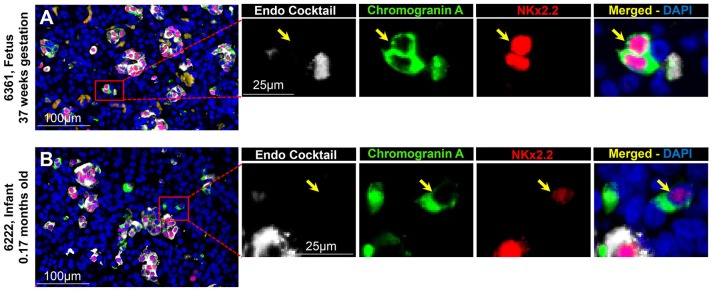
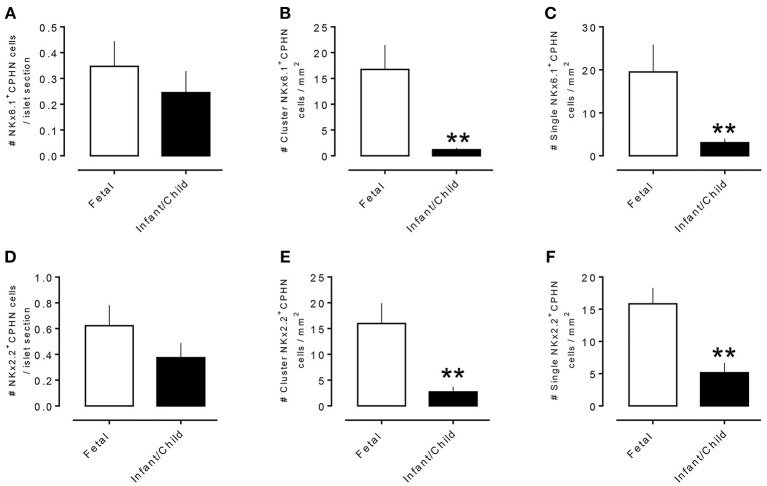
Context: Previously, we identified chromograninA positive hormone-negative (CPHN) cells in high frequency in human fetal and neonatal pancreas, likely representing nascent endocrine precursor cells. Here, we characterize the putative endocrine fate and replicative status of these newly formed cells. Objective: To establish the replicative frequency and transcriptional identity of CPHN cells, extending our observation on CPHN cell frequency to a larger cohort of fetal and infant pancreas. Design, Setting, and Participants: 8 fetal, 19 infant autopsy pancreata were evaluated for CPHN cell frequency; 12 fetal, 24 infant/child pancreata were evaluated for CPHN replication and identity. Results: CPHN cell frequency decreased 84% (islets) and 42% (clusters) from fetal to infant life. Unlike the beta-cells at this stage, CPHN cells were rarely observed to replicate (0.2 ± 0.1 vs. 4.7 ± 1.0%, CPHN vs. islet hormone positive cell replication, p < 0.001), indicated by the lack of Ki67 expression in CPHN cells whether located in the islets or in small clusters, and with no detectable difference between fetal and infant groups. While the majority of CPHN cells express (in overall compartments of pancreas) the pan-endocrine transcription factor NKX2.2 and beta-cell specific NKX6.1 in comparable frequency in fetal and infant/child cases (81.9 ± 6.3 vs. 82.8 ± 3.8% NKX6.1+-CPHN cells of total CPHN cells, fetal vs. infant/child, p = 0.9; 88.0 ± 4.7 vs. 82.1 ± 5.3% NKX2.2+-CPHN cells of total CPHN cells, fetal vs. infant/child, p = 0.4), the frequency of clustered CPHN cells expressing NKX6.1 or NKX2.2 is lower in infant/child vs. fetal cases (1.2 ± 0.3 vs. 16.7 ± 4.7 clustered NKX6.1+-CPHN cells/mm2, infant/child vs. fetal, p < 0.01; 2.7 ± 1.0 vs. 16.0 ± 4.0 clustered NKX2.2+-CPHN cells/mm2, infant/child vs. fetal, p < 0.01). Conclusions: The frequency of CPHN cells declines steeply from fetal to infant life, presumably as they differentiate to hormone-expressing cells. CPHN cells represent a non-replicative pool of endocrine precursor cells, a proportion of which are likely fated to become beta-cells. Precis : CPHN cell frequency declines steeply from fetal to infant life, as they mature to hormone expression. CPHN cells represent a non-replicative pool of endocrine precursor cells, a proportion of which are likely fated to become beta-cells.
Keywords: beta-cell; development; endocrine cells; maturation; replication; transcription factor.
Figures


References
LinkOut - more resources
Full Text Sources