

Epicatechin modulates stress-resistance in C. elegans via insulin/IGF-1 signaling pathway
- PMID: 30689636
- PMCID: PMC6349306
- DOI: 10.1371/journal.pone.0199483
Epicatechin modulates stress-resistance in C. elegans via insulin/IGF-1 signaling pathway
Abstract
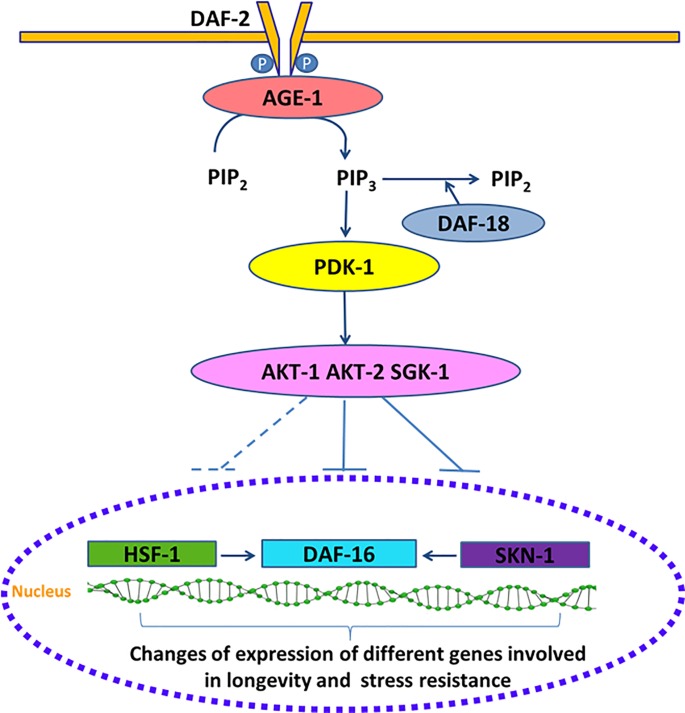
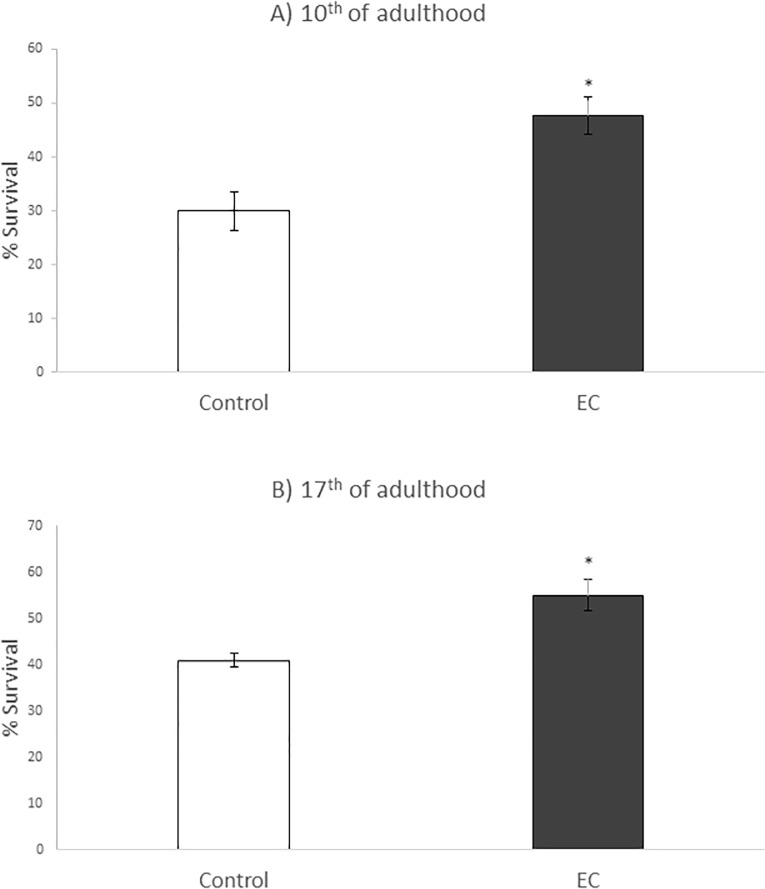
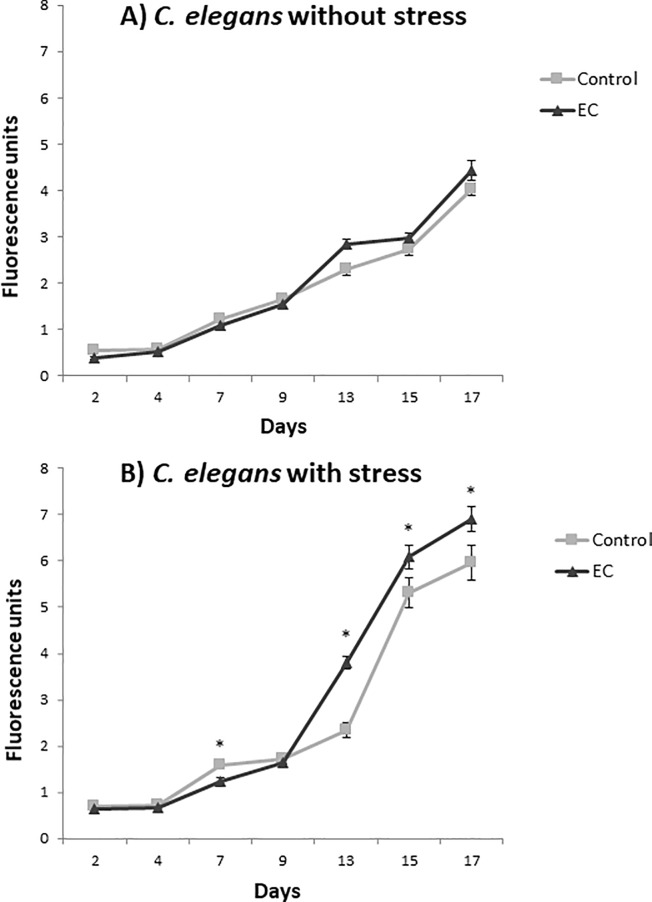
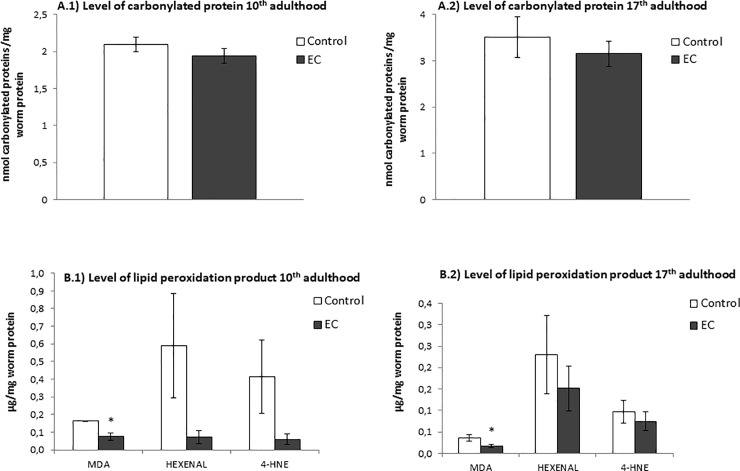
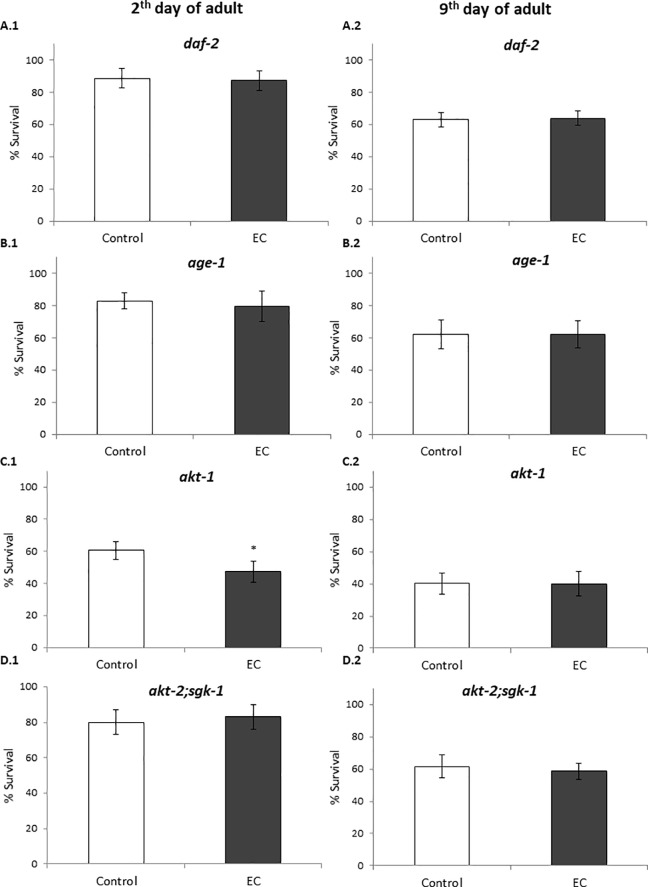
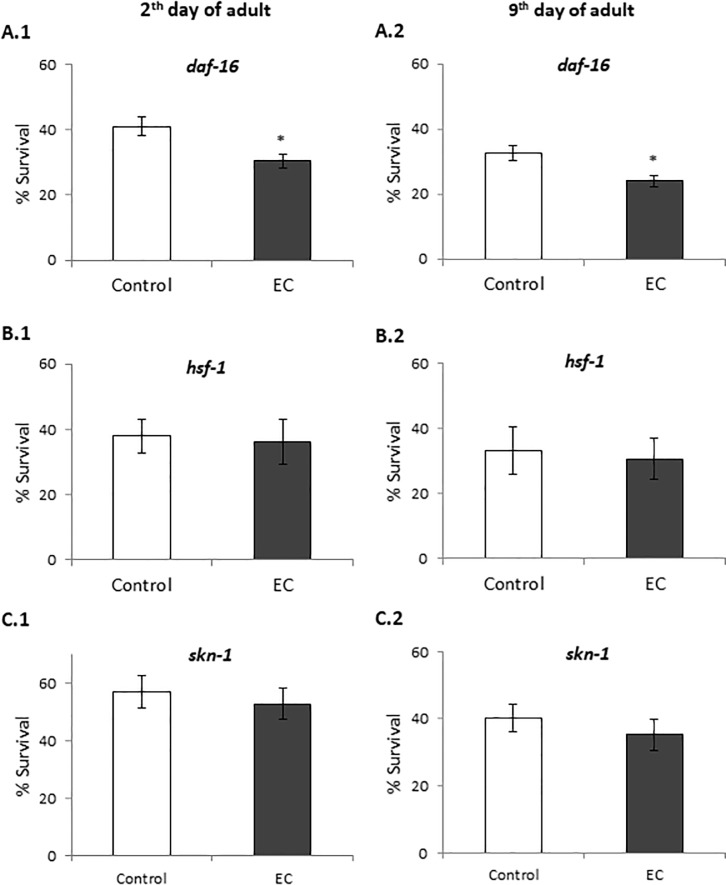
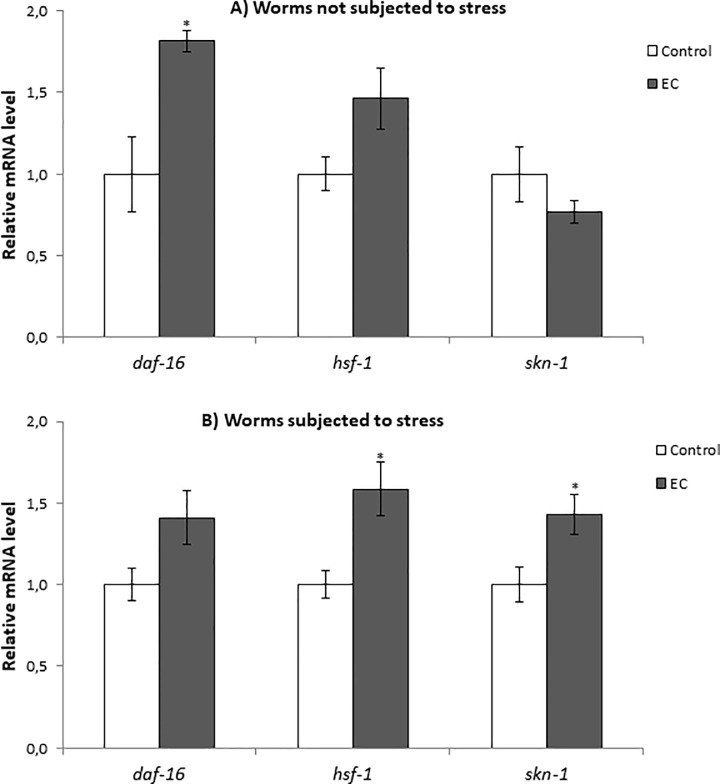
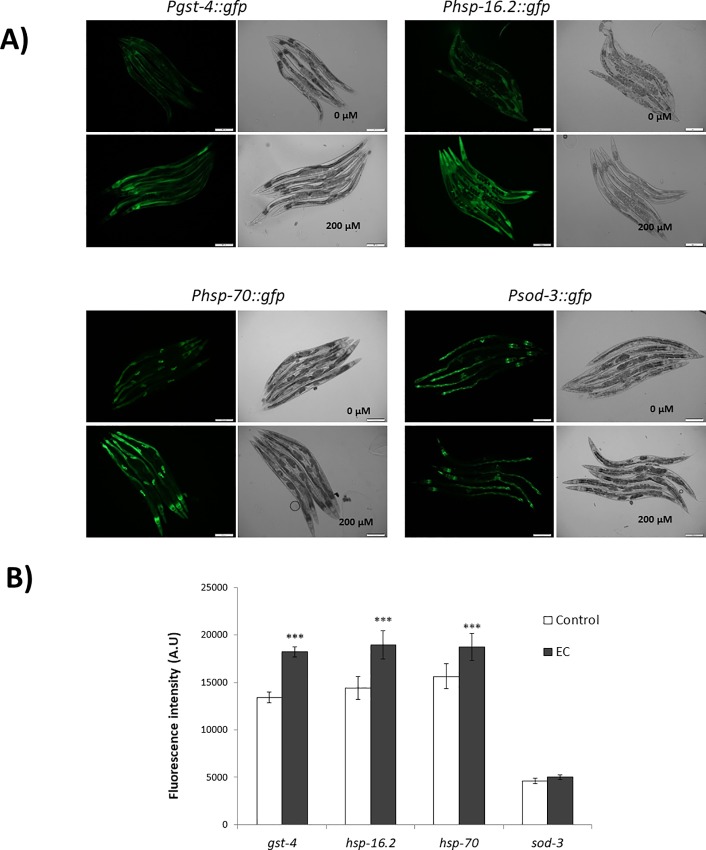
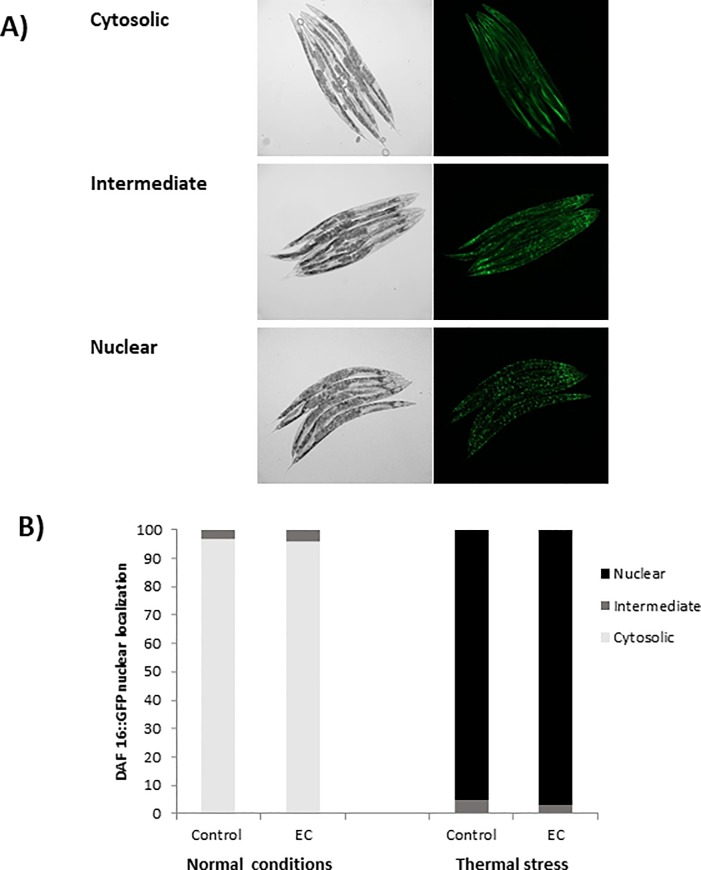
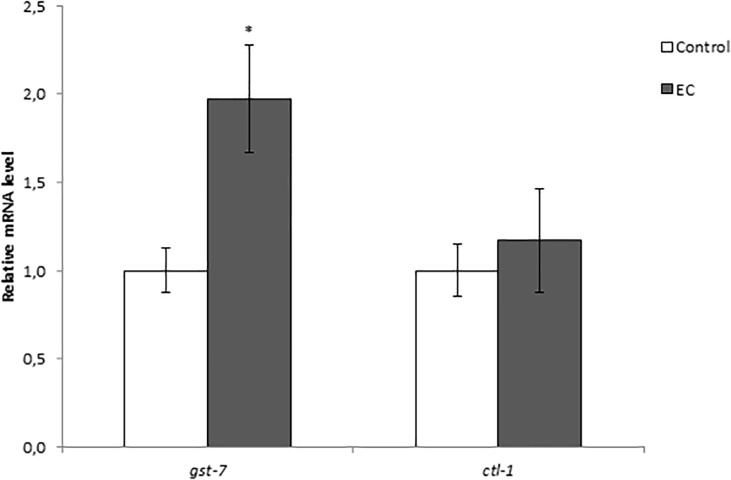
The nematode Caenorhabditis elegans has been used to examine the influence of epicatechin (EC), an abundant flavonoid in the human diet, in some stress biomarkers (ROS production, lipid peroxidation and protein carbonylation). Furthermore, the ability of EC to modulate the expression of some key genes in the insulin/IGF-1 signaling pathway (IIS), involved in longevity and oxidative or heat shock stress response, has also been explored. The final aim was to contribute to the elucidation of the mechanisms involved in the biological effects of flavonoids. The results showed that EC-treated wild-type C. elegans exhibited increased survival and reduced oxidative damage of biomolecules when submitted to thermal stress. EC treatment led to a moderate elevation in ROS levels, which might activate endogenous mechanisms of defense protecting against oxidative insult. The enhanced stress resistance induced by EC was found to be mediated through the IIS pathway, since assays in daf-2, age-1, akt-1, akt-2, sgk-1, daf-16, skn-1 and hsf-1 loss of function mutant strains failed to show any heat-resistant phenotype against thermal stress when treated with EC. Consistently, EC treatment upregulated the expression of some stress resistance associated genes, such as gst-4, hsp-16.2 and hsp-70, which are downstream regulated by the IIS pathway.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Santos-Buelga C, Scalbert A. Proanthocyanidins and tannin‐like compounds–nature, occurrence, dietary intake and effects on nutrition and health. J. Sci. Food Agric. 2000;80: 1094–1117.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
