

Reduction of the expression of the late-onset Alzheimer's disease (AD) risk-factor BIN1 does not affect amyloid pathology in an AD mouse model
- PMID: 30692199
- PMCID: PMC6433054
- DOI: 10.1074/jbc.RA118.006379
Reduction of the expression of the late-onset Alzheimer's disease (AD) risk-factor BIN1 does not affect amyloid pathology in an AD mouse model
Abstract
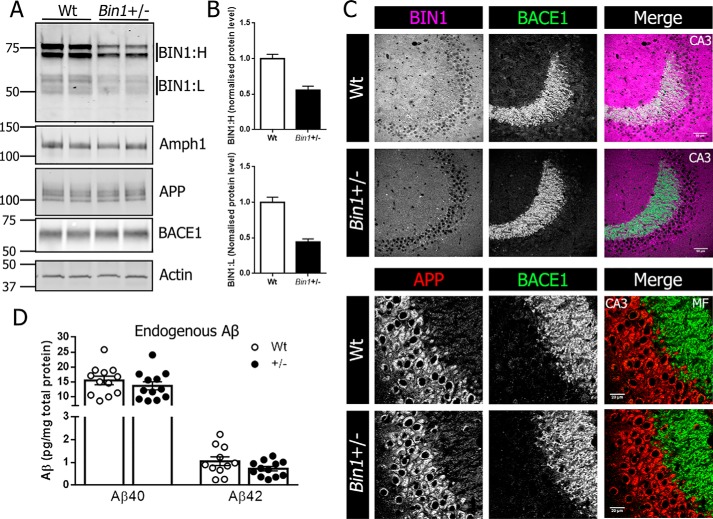
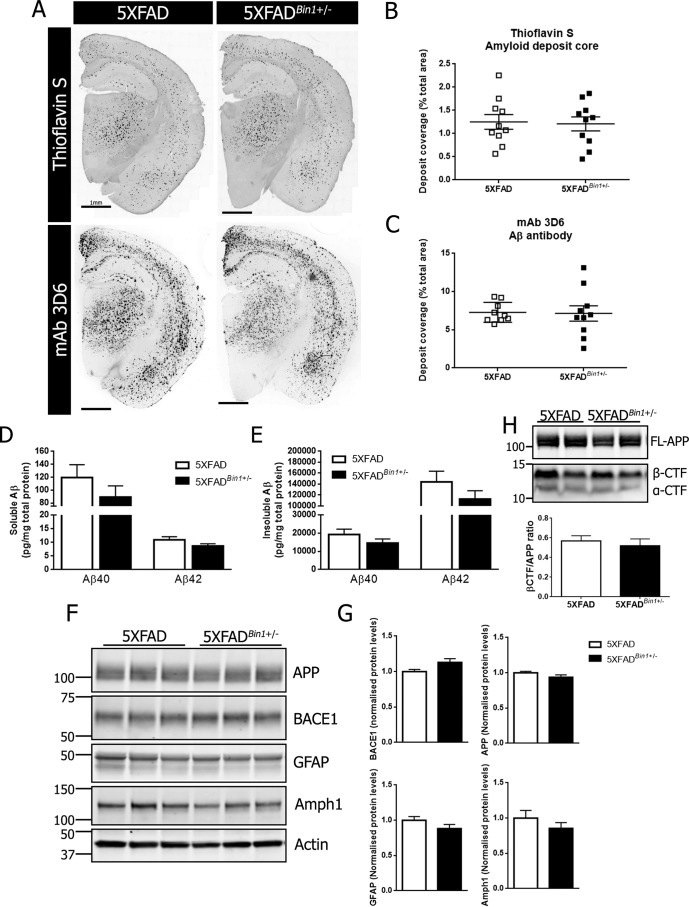
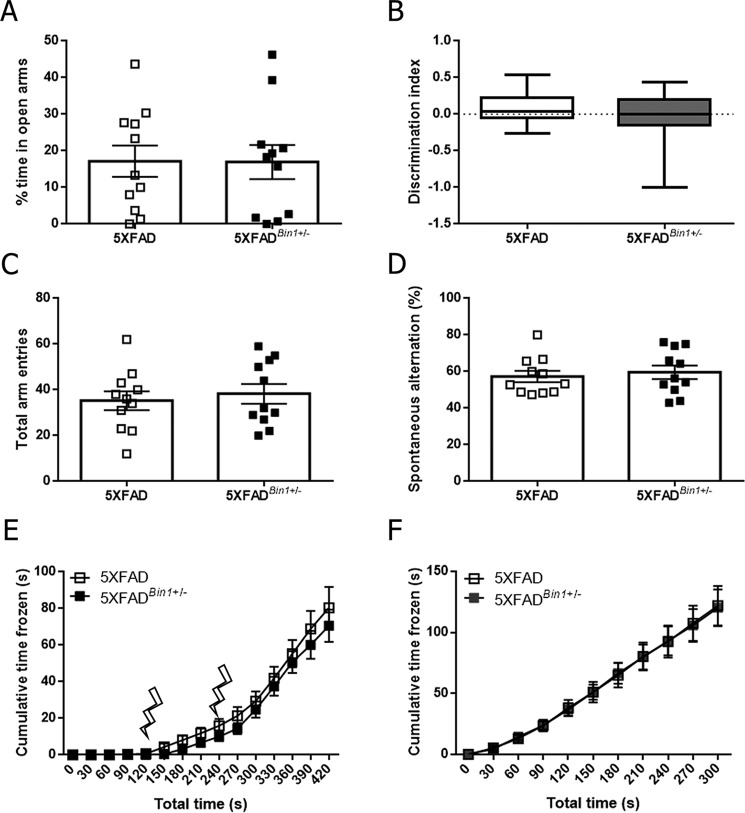
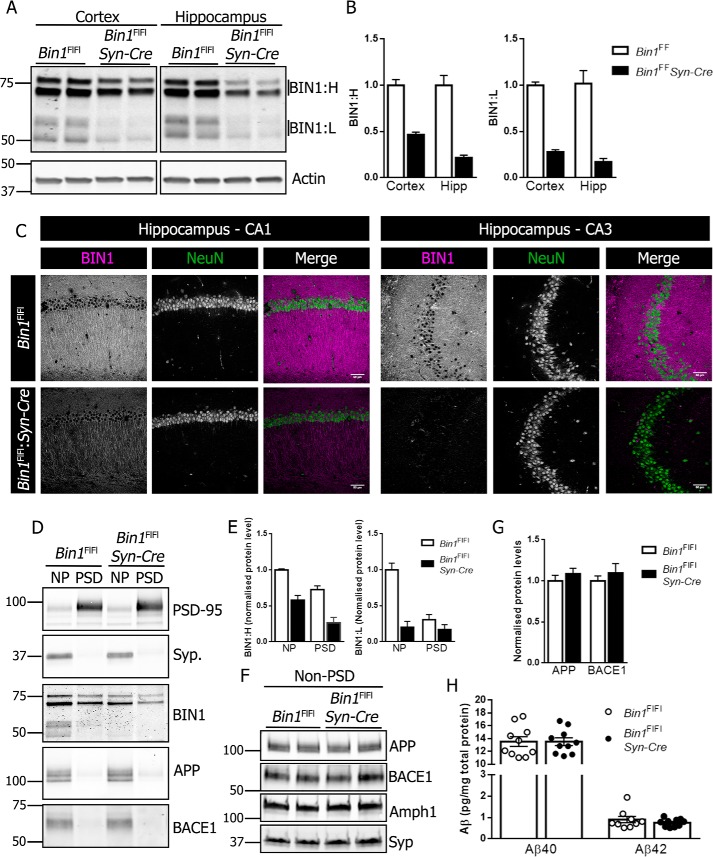
Alzheimer's disease (AD) is pathologically characterized by the deposition of the β-amyloid (Aβ) peptide in senile plaques in the brain, leading to neuronal dysfunction and eventual decline in cognitive function. Genome-wide association studies have identified the bridging integrator 1 (BIN1) gene within the second most significant susceptibility locus for late-onset AD. BIN1 is a member of the amphiphysin family of proteins and has reported roles in the generation of membrane curvature and endocytosis. Endocytic dysfunction is a pathological feature of AD, and endocytosis of the amyloid precursor protein is an important step in its subsequent cleavage by β-secretase (BACE1). In vitro evidence implicates BIN1 in endosomal sorting of BACE1 and Aβ generation in neurons, but a role for BIN1 in this process in vivo is yet to be described. Here, using biochemical and immunohistochemistry analyses we report that a 50% global reduction of BIN1 protein levels resulting from a single Bin1 allele deletion in mice does not change BACE1 levels or localization in vivo, nor does this reduction alter the production of endogenous murine Aβ in nontransgenic mice. Furthermore, we found that reduction of BIN1 levels in the 5XFAD mouse model of amyloidosis does not alter Aβ deposition nor behavioral deficits associated with cerebral amyloid burden. Finally, a conditional BIN1 knockout in excitatory neurons did not alter BACE1, APP, C-terminal fragments derived from BACE1 cleavage of APP, or endogenous Aβ levels. These results indicate that BIN1 function does not regulate Aβ generation in vivo.
Keywords: 5XFAD; APP; Alzheimer disease; BACE1; BIN1; GWAS; amphiphysin; amyloid; amyloid precursor protein (APP); beta-amyloid (AB); beta-secretase 1 (BACE1); endocytosis; neurodegeneration; pathogenesis; secretase.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Harold D., Abraham R., Hollingworth P., Sims R., Gerrish A., Hamshere M. L., Pahwa J. S., Moskvina V., Dowzell K., Williams A., Jones N., Thomas C., Stretton A., Morgan A. R., Lovestone S., et al. (2009) Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer's disease. Nat. Genet. 41, 1088–1093 10.1038/ng.440 - DOI - PMC - PubMed
-
- Naj A. C., Jun G., Beecham G. W., Wang L.-S., Vardarajan B. N., Buros J., Gallins P. J., Buxbaum J. D., Jarvik G. P., Crane P. K., Larson E. B., Bird T. D., Boeve B. F., Graff-Radford N. R., De Jager P. L., et al. (2011) Common variants at MS4A4/MS4A6E, CD2AP, CD33 and EPHA1 are associated with late-onset Alzheimer's disease. Nat. Genet. 43, 436–441 10.1038/ng.801 - DOI - PMC - PubMed
-
- Seshadri S., Fitzpatrick A. L., Ikram M. A., DeStefano A. L., Gudnason V., Boada M., Bis J. C., Smith A. V., Carassquillo M. M., Lambert J. C., Harold D., Schrijvers E. M., Ramirez-Lorca R., Debette S., Longstreth W. T. Jr., et al. (2010) Genome-wide analysis of genetic loci associated with Alzheimer disease. JAMA 303, 1832–1840 10.1001/jama.2010.574 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
