

CD44 splice isoform switching determines breast cancer stem cell state
- PMID: 30692202
- PMCID: PMC6362815
- DOI: 10.1101/gad.319889.118
CD44 splice isoform switching determines breast cancer stem cell state
Abstract
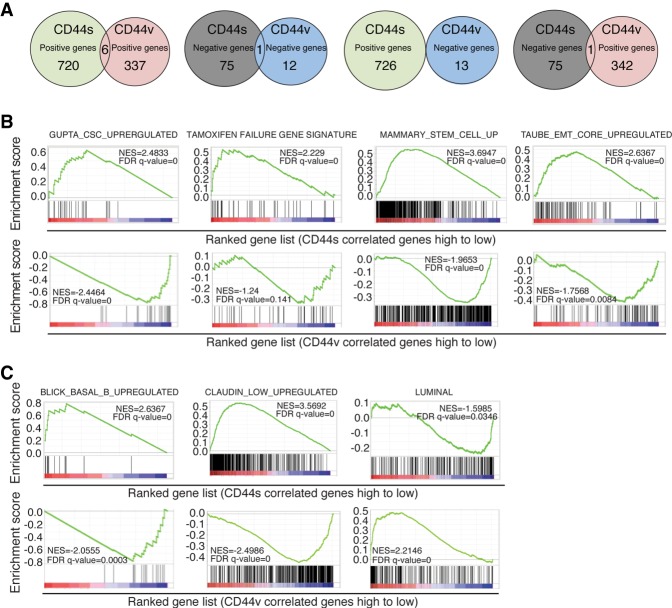
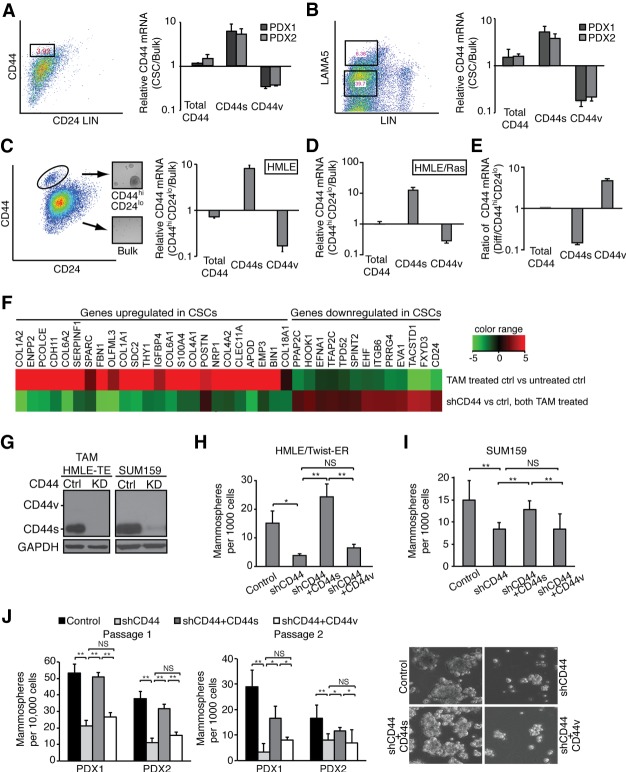
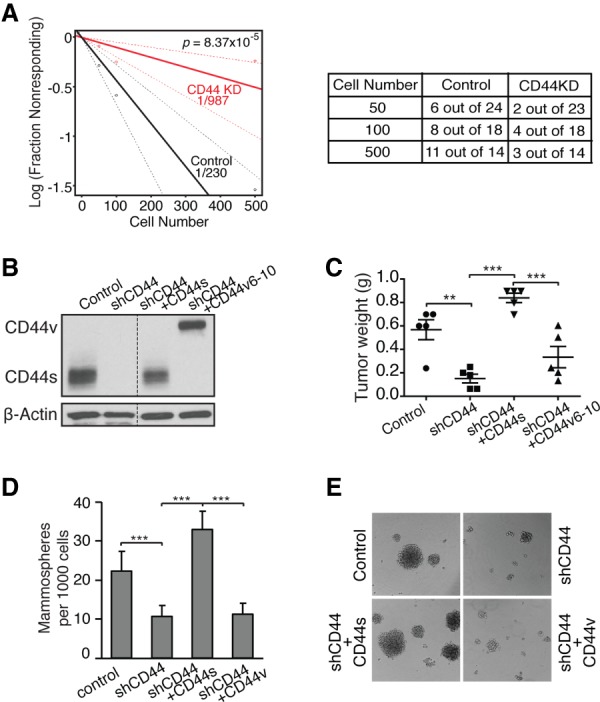
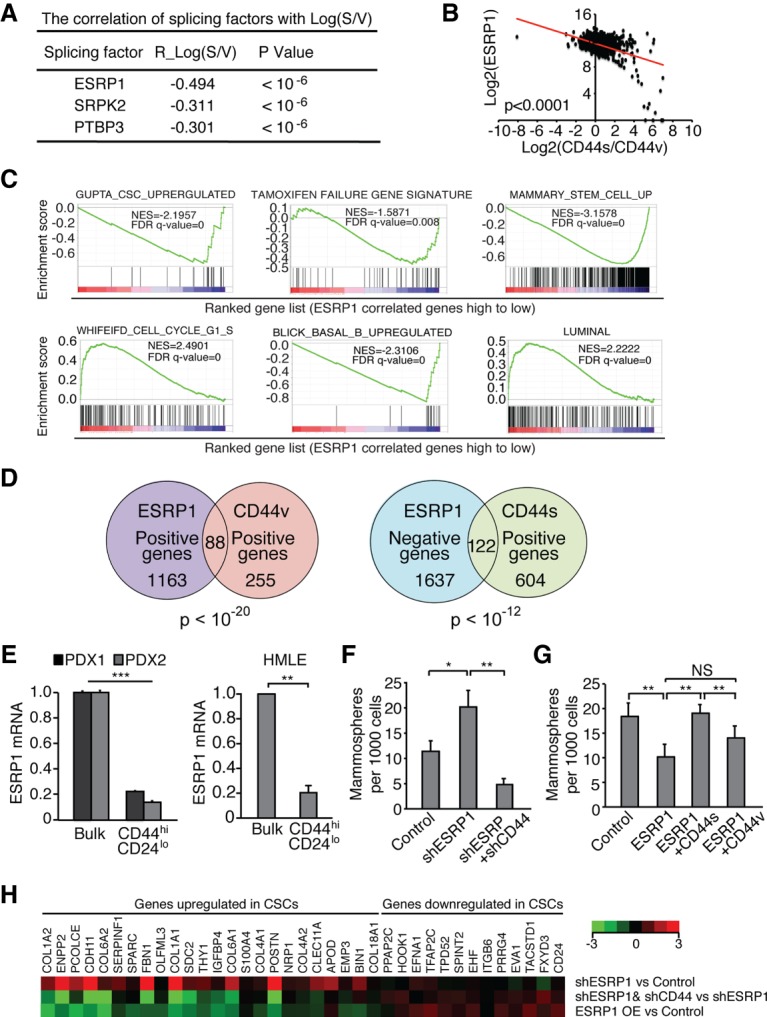
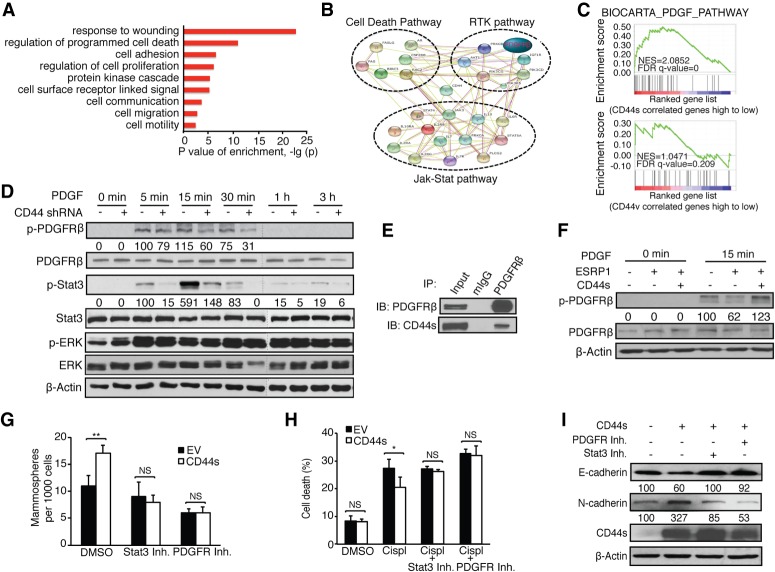
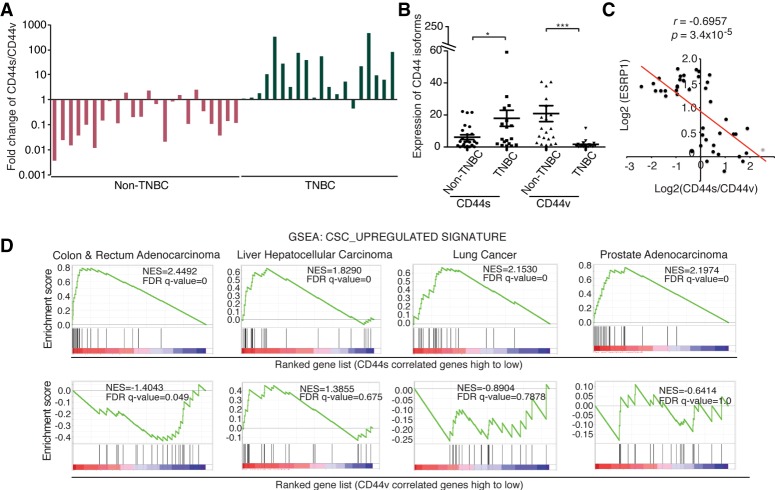
Although changes in alternative splicing have been observed in cancer, their functional contributions still remain largely unclear. Here we report that splice isoforms of the cancer stem cell (CSC) marker CD44 exhibit strikingly opposite functions in breast cancer. Bioinformatic annotation in patient breast cancer in The Cancer Genome Atlas (TCGA) database reveals that the CD44 standard splice isoform (CD44s) positively associates with the CSC gene signatures, whereas the CD44 variant splice isoforms (CD44v) exhibit an inverse association. We show that CD44s is the predominant isoform expressed in breast CSCs. Elimination of the CD44s isoform impairs CSC traits. Conversely, manipulating the splicing regulator ESRP1 to shift alternative splicing from CD44v to CD44s leads to an induction of CSC properties. We further demonstrate that CD44s activates the PDGFRβ/Stat3 cascade to promote CSC traits. These results reveal CD44 isoform specificity in CSC and non-CSC states and suggest that alternative splicing provides functional gene versatility that is essential for distinct cancer cell states and thus cancer phenotypes.
Keywords: CD44s; CSC; PDGFRβ/Stat3; alternative splicing; breast cancer.
© 2019 Zhang et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous