

Nuclear pore density controls heterochromatin reorganization during senescence
- PMID: 30692205
- PMCID: PMC6362808
- DOI: 10.1101/gad.321117.118
Nuclear pore density controls heterochromatin reorganization during senescence
Abstract
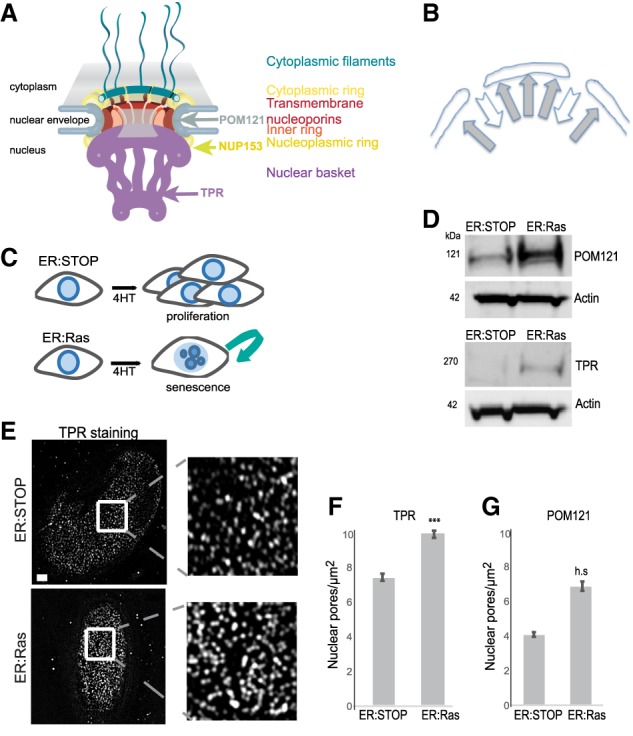
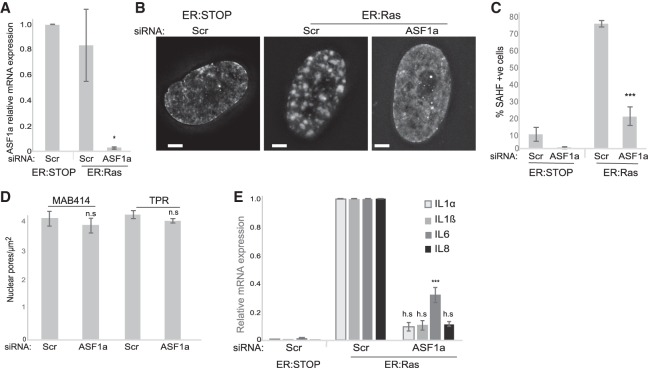
During oncogene-induced senescence (OIS), heterochromatin is lost from the nuclear periphery and forms internal senescence-associated heterochromatin foci (SAHFs). We show that an increased nuclear pore density during OIS is responsible for SAHF formation. In particular, the nucleoporin TPR is necessary for both formation and maintenance of SAHFs. Loss of SAHFs does not affect cell cycle arrest but abrogates the senescence-associated secretory phenotype-a program of inflammatory cytokine gene activation. Our results uncover a previously unknown role of nuclear pores in heterochromatin reorganization in mammalian nuclei and demonstrate the importance of heterochromatin organization for a specific gene activation program.
Keywords: inflammation; nuclear organization; nuclear pore; senescence.
© 2019 Boumendil et al.; Published by Cold Spring Harbor Laboratory Press.
Figures



Comment in
-
The unusual SASPects.Nat Rev Mol Cell Biol. 2019 Apr;20(4):195. doi: 10.1038/s41580-019-0111-9. Nat Rev Mol Cell Biol. 2019. PMID: 30770903 No abstract available.
References
-
- Bodoor K, Shaikh S, Salina D, Raharjo WH, Bastos R, Lohka M, Burke B. 1999. Sequential recruitment of NPC proteins to the nuclear periphery at the end of mitosis. J Cell Sci 112: 2253–2264. - PubMed
-
- Chandra T, Kirschner K, Thuret J-Y, Pope BD, Ryba T, Newman S, Ahmed K, Samarajiwa SA, Salama R, Carroll T, et al. 2012. Independence of repressive histone marks and chromatin compaction during senescent heterochromatic layer formation. Mol Cell 47: 203–214. 10.1016/j.molcel.2012.06.010 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources