

Dominance reversals and the maintenance of genetic variation for fitness
- PMID: 30695026
- PMCID: PMC6368311
- DOI: 10.1371/journal.pbio.3000118
Dominance reversals and the maintenance of genetic variation for fitness
Abstract
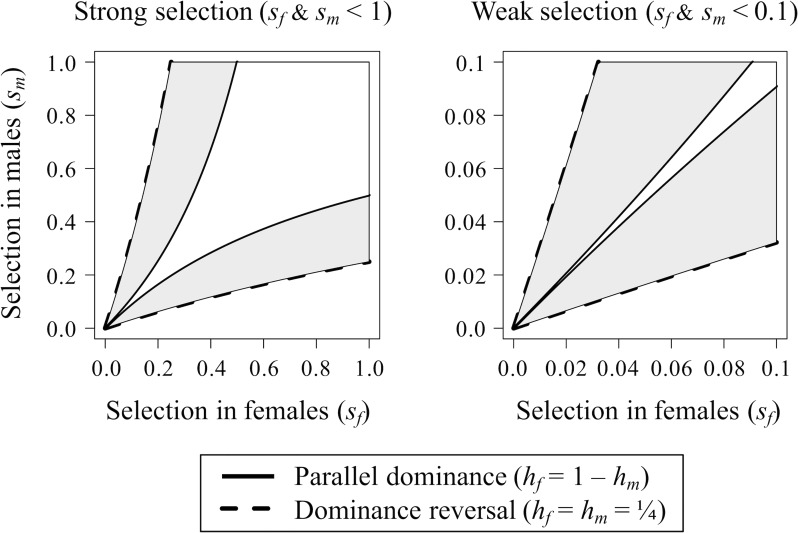
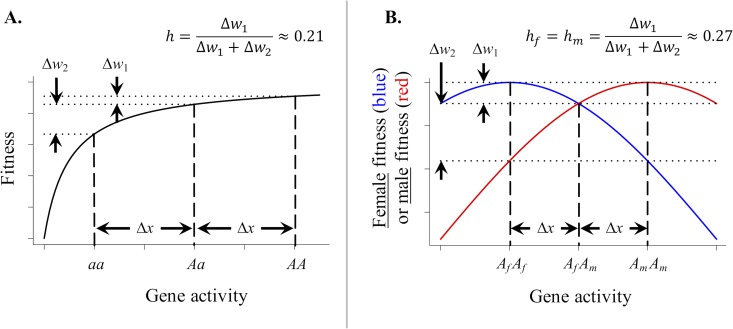
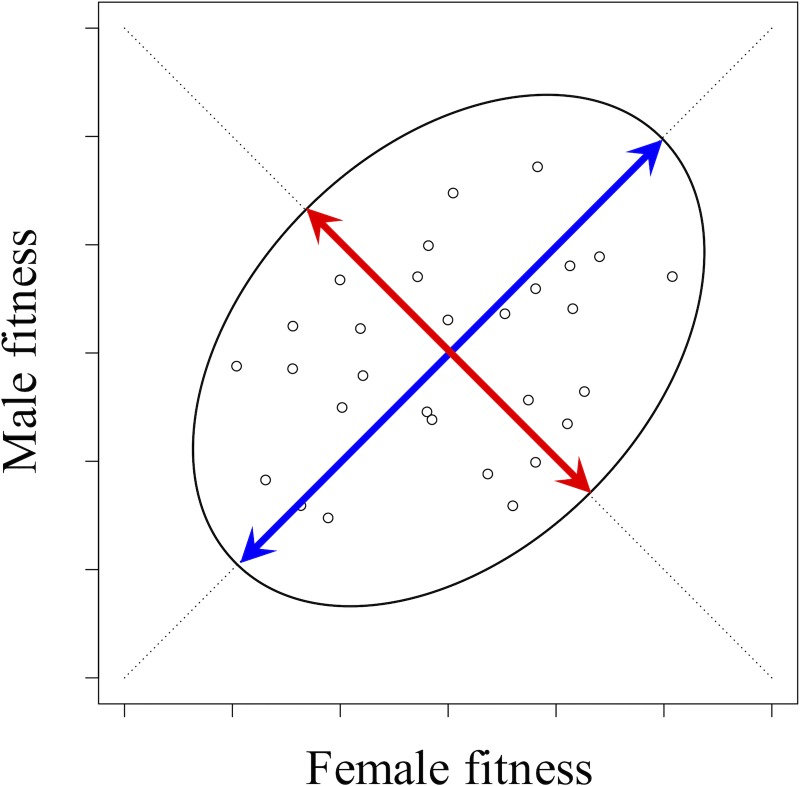
Antagonistic selection between different fitness components (e.g., survival versus fertility) or different types of individuals in a population (e.g., females versus males) can potentially maintain genetic diversity and thereby account for the high levels of fitness variation observed in natural populations. However, the degree to which antagonistic selection can maintain genetic variation critically depends on the dominance relations between antagonistically selected alleles in diploid individuals. Conditions for stable polymorphism of antagonistically selected alleles are narrow, particularly when selection is weak, unless the alleles exhibit "dominance reversals"-in which each allele is partially or completely dominant in selective contexts in which it is favored and recessive in contexts in which it is harmful. Although theory predicts that dominance reversals should emerge under biologically plausible conditions, evidence for dominance reversals is sparse. In this primer, we review theoretical arguments and data supporting a role for dominance reversals in the maintenance of genetic variation. We then highlight an illuminating new study by Grieshop and Arnqvist, which reports a genome-wide signal of dominance reversals between male and female fitness in seed beetles.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment on
-
Sex-specific dominance reversal of genetic variation for fitness.PLoS Biol. 2018 Dec 11;16(12):e2006810. doi: 10.1371/journal.pbio.2006810. eCollection 2018 Dec. PLoS Biol. 2018. PMID: 30533008 Free PMC article.
References
-
- Lynch M, Walsh B. Genetics and analysis of quantitative traits. Sunderland: Sinauer Associates; 1998.
-
- Walsh B, Blows MW. Abundant genetic variation + strong selection = multivariate genetic constraints: a geometric view of adaptation. Ann Rev Ecol Evol Syst. 2009;40: 41–59.
-
- Dobzhansky T. A review of some fundamental concepts and problems of population genetics. Cold Spring Harbor Symposia Quant Biol. 1955;20: 1–15. - PubMed
-
- Lewontin RC. The genetic basis of evolutionary change. New York: Columbia University Press; 1974.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
