

Functional characterization of the alanine-serine-cysteine exchanger of Carnobacterium sp AT7
- PMID: 30696726
- PMCID: PMC6445583
- DOI: 10.1085/jgp.201812195
Functional characterization of the alanine-serine-cysteine exchanger of Carnobacterium sp AT7
Abstract
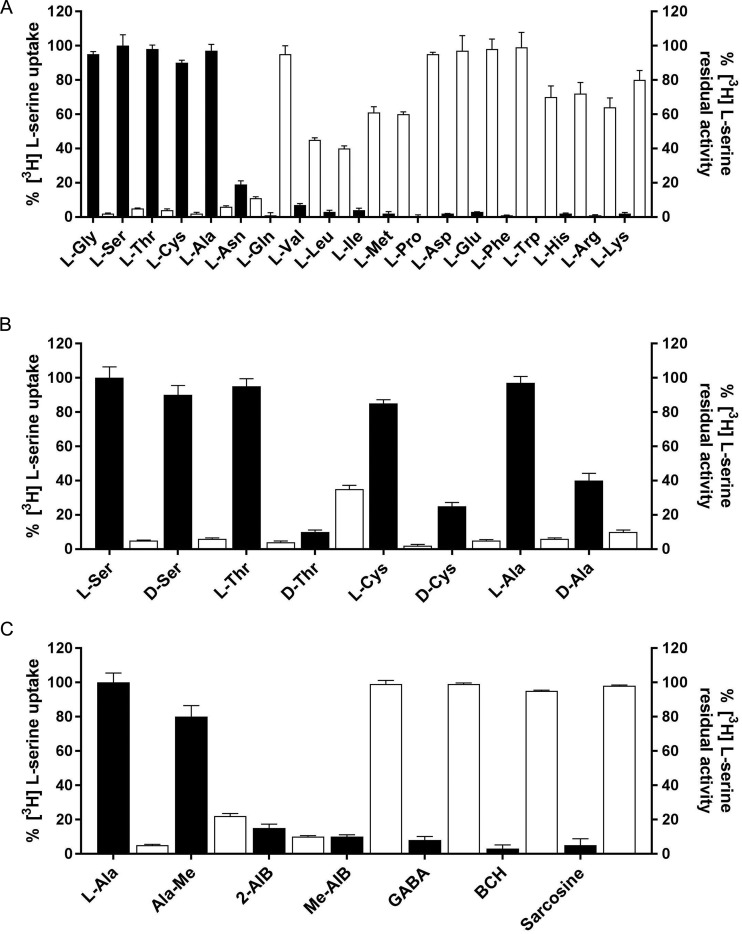
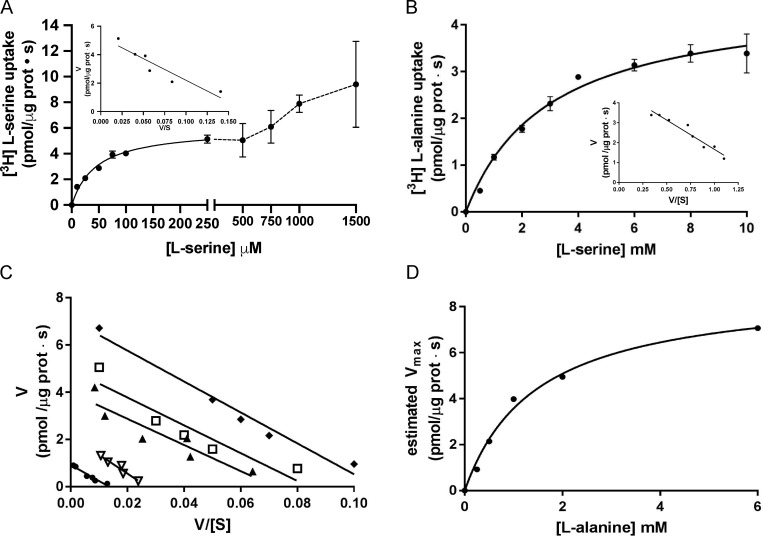
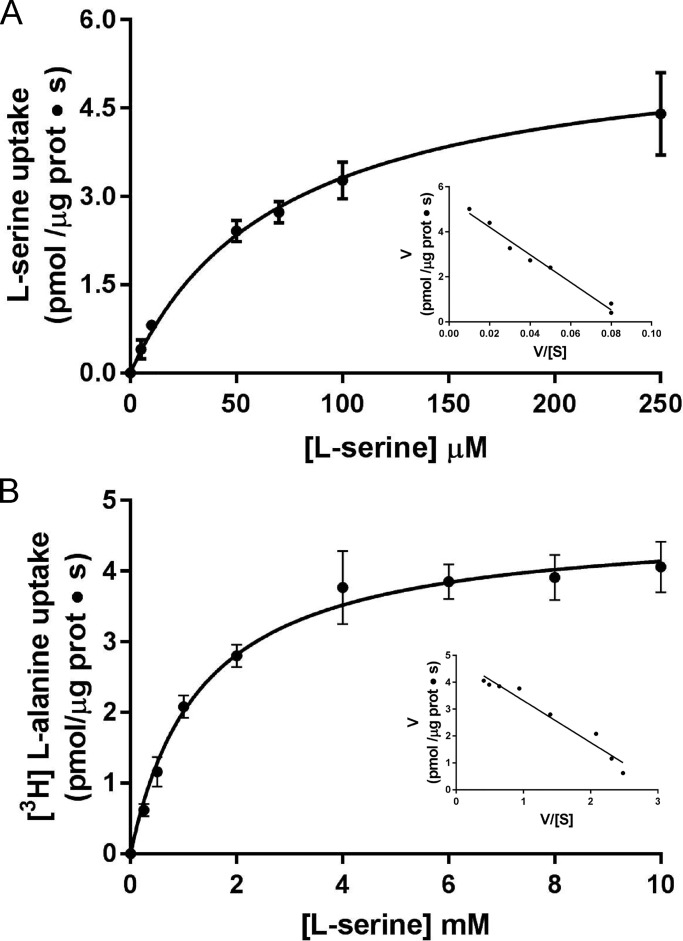
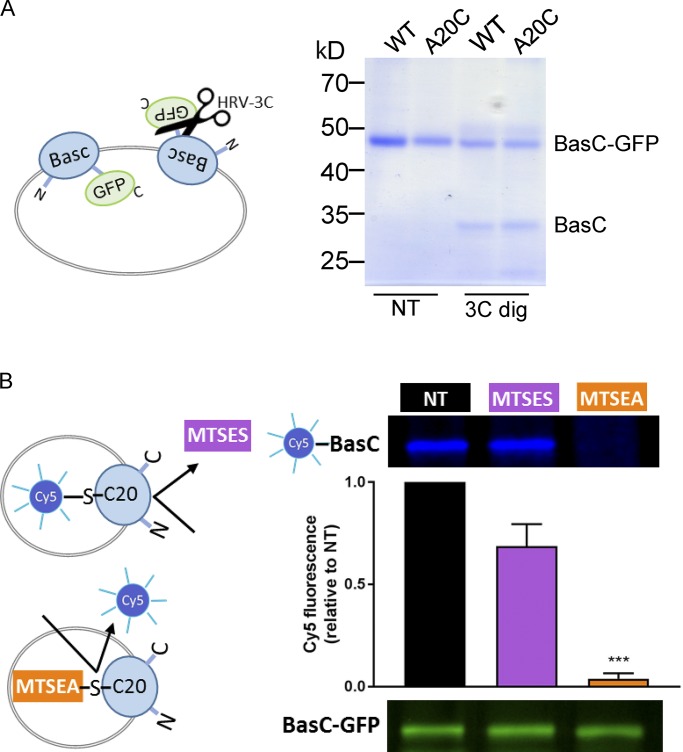
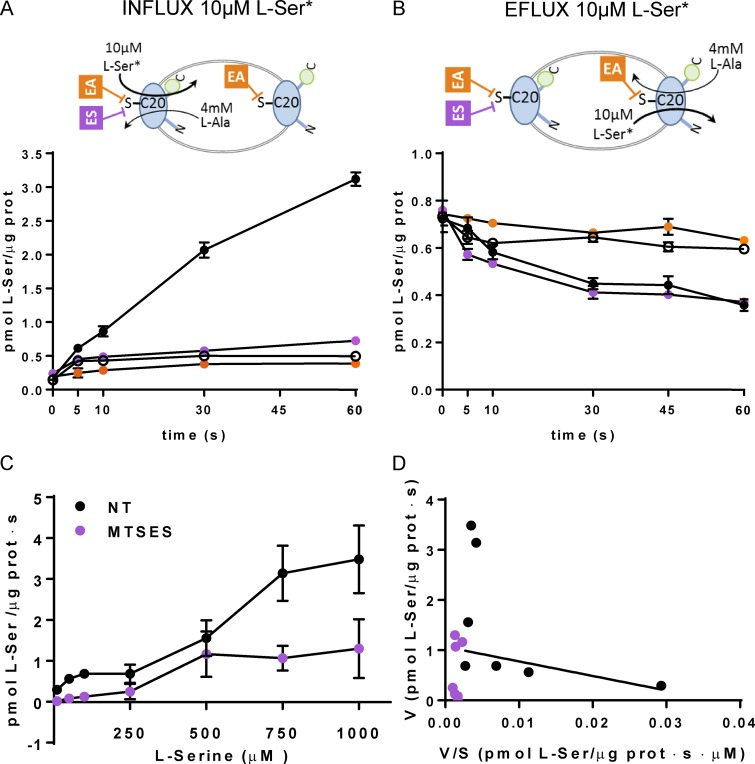
Many key cell processes require prior cell uptake of amino acids from the environment, which is facilitated by cell membrane amino acid transporters such as those of the L-type amino acid transporter (LAT) subfamily. Alterations in LAT subfamily amino acid transport are associated with several human diseases, including cancer, aminoacidurias, and neurodegenerative conditions. Therefore, from the perspective of human health, there is considerable interest in obtaining structural information about these transporter proteins. We recently solved the crystal structure of the first LAT transporter, the bacterial alanine-serine-cysteine exchanger of Carnobacterium sp AT7 (BasC). Here, we provide a complete functional characterization of detergent-purified, liposome-reconstituted BasC transporter to allow the extension of the structural insights into mechanistic understanding. BasC is a sodium- and proton-independent small neutral amino acid exchanger whose substrate and inhibitor selectivity are almost identical to those previously described for the human LAT subfamily member Asc-1. Additionally, we show that, like its human counterparts, this transporter has apparent affinity asymmetry for the intra- and extracellular substrate binding sites-a key feature in the physiological role played by these proteins. BasC is an excellent paradigm of human LAT transporters and will contribute to our understanding of the molecular mechanisms underlying substrate recognition and translocation at both sides of the plasma membrane.
© 2019 Bartoccioni et al.
Figures
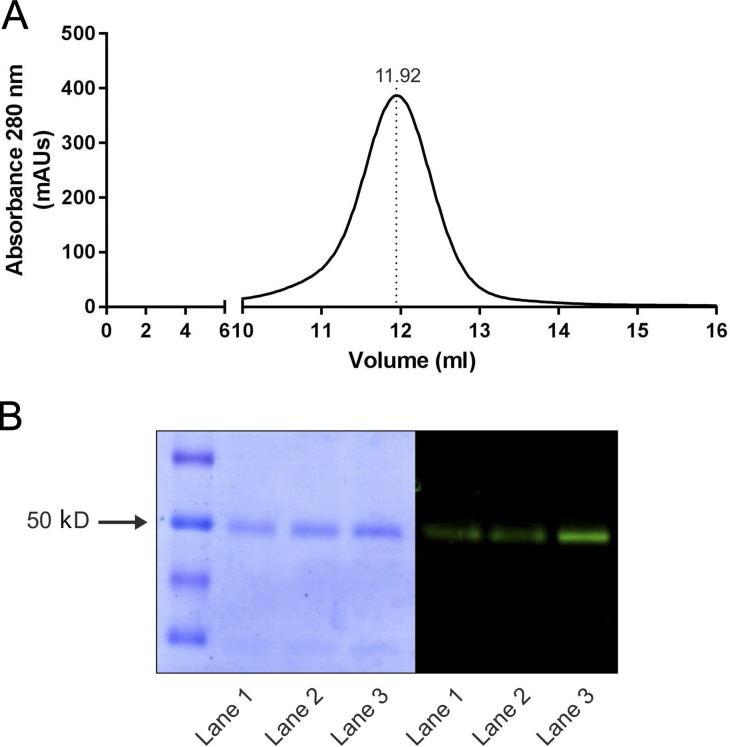
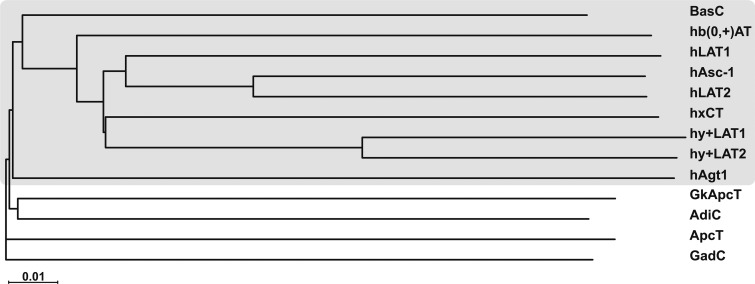
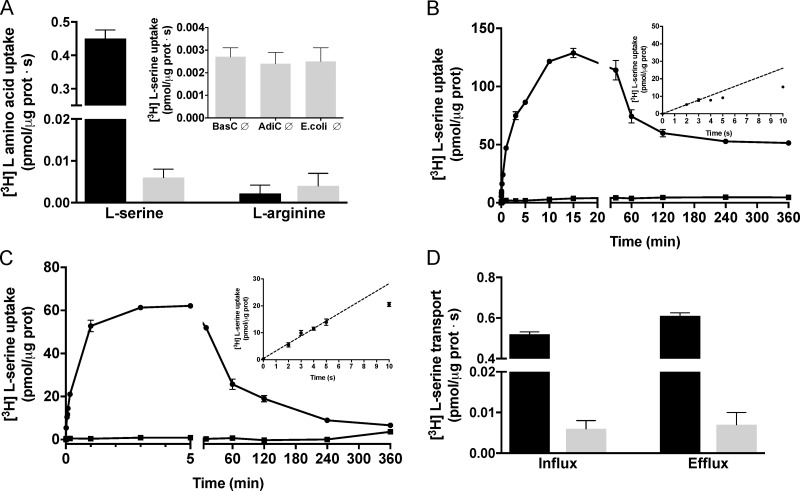
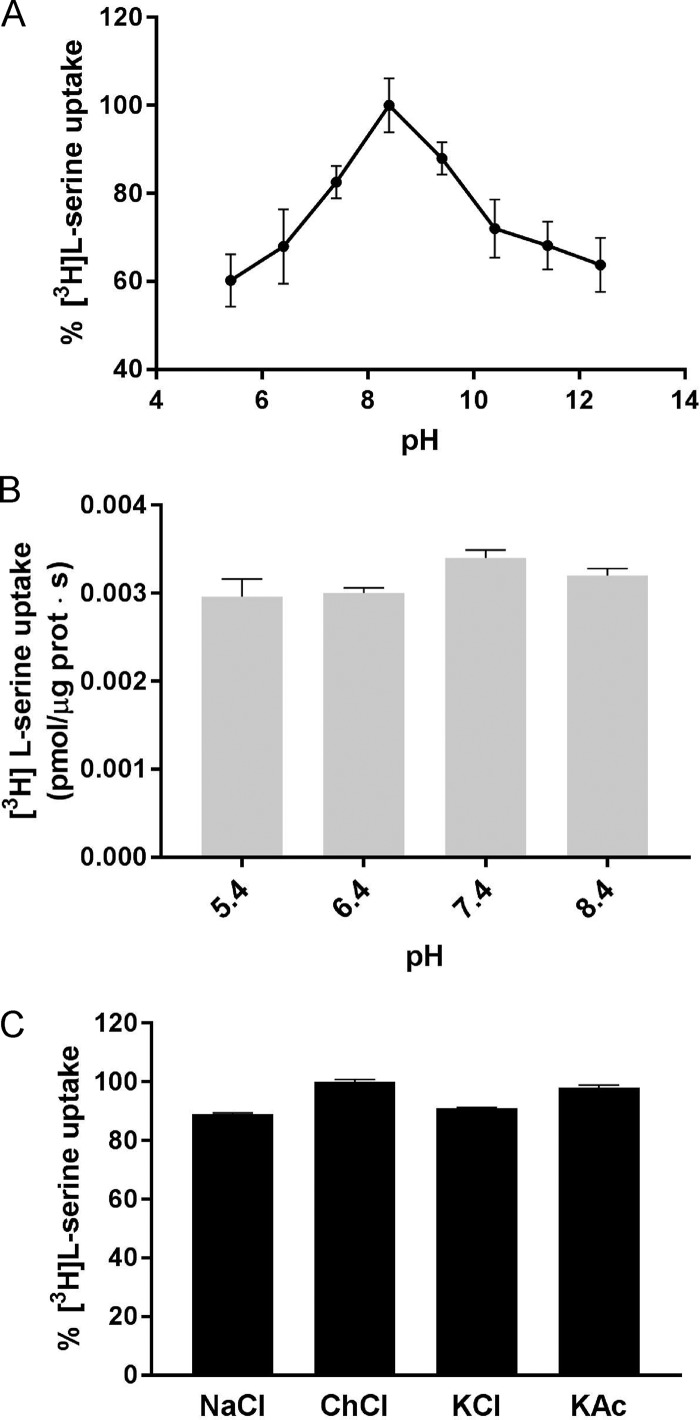
References
-
- Borsani G., Bassi M.T., Sperandeo M.P., De Grandi A., Buoninconti A., Riboni M., Manzoni M., Incerti B., Pepe A., Andria G., et al. 1999. SLC7A7, encoding a putative permease-related protein, is mutated in patients with lysinuric protein intolerance. Nat. Genet. 21:297–301. 10.1038/6815 - DOI - PubMed
-
- Brown J.M., Hunihan L., Prack M.M., Harden D.G., Bronson J., Dzierba C.D., Gentles R.G., Hendricson A., Krause R., Macor J.E., and Westphal R.S.. 2014. In vitro Characterization of a small molecule inhibitor of the alanine serine cysteine transporter -1 (SLC7A10). J. Neurochem. 129:275–283. 10.1111/jnc.12618 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
