

Theta/delta coupling across cortical laminae contributes to semantic cognition
- PMID: 30699059
- PMCID: PMC6485732
- DOI: 10.1152/jn.00686.2018
Theta/delta coupling across cortical laminae contributes to semantic cognition
Abstract
Rhythmic activity in populations of neurons is associated with cognitive and motor function. Our understanding of the neuronal mechanisms underlying these core brain functions has benefitted from demonstrations of cellular, synaptic, and network phenomena, leading to the generation of discrete rhythms at the local network level. However, discrete frequencies of rhythmic activity rarely occur alone. Despite this, little is known about why multiple rhythms are generated together or what mechanisms underlie their interaction to promote brain function. One overarching theory is that different temporal scales of rhythmic activity correspond to communication between brain regions separated by different spatial scales. To test this, we quantified the cross-frequency interactions between two dominant rhythms-theta and delta activity-manifested during magnetoencephalography recordings of subjects performing a word-pair semantic decision task. Semantic processing has been suggested to involve the formation of functional links between anatomically disparate neuronal populations over a range of spatial scales, and a distributed network was manifest in the profile of theta-delta coupling seen. Furthermore, differences in the pattern of theta-delta coupling significantly correlated with semantic outcome. Using an established experimental model of concurrent delta and theta rhythms in neocortex, we show that these outcome-dependent dynamics could be reproduced in a manner determined by the strength of cholinergic neuromodulation. Theta-delta coupling correlated with discrete neuronal activity motifs segregated by the cortical layer, neuronal intrinsic properties, and long-range axonal targets. Thus, the model suggested that local, interlaminar neocortical theta-delta coupling may serve to coordinate both cortico-cortical and cortico-subcortical computations during distributed network activity. NEW & NOTEWORTHY Here, we show, for the first time, that a network of spatially distributed brain regions can be revealed by cross-frequency coupling between delta and theta frequencies in subjects using magnetoencephalography recording during a semantic decision task. A biological model of this cross-frequency coupling suggested an interlaminar, cell-specific division of labor within the neocortex may serve to route the flow of cortico-cortical and cortico-subcortical information to promote such spatially distributed, functional networks.
Keywords: cross-frequency coupling; delta rhythm; semantic processing; theta rhythm.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures
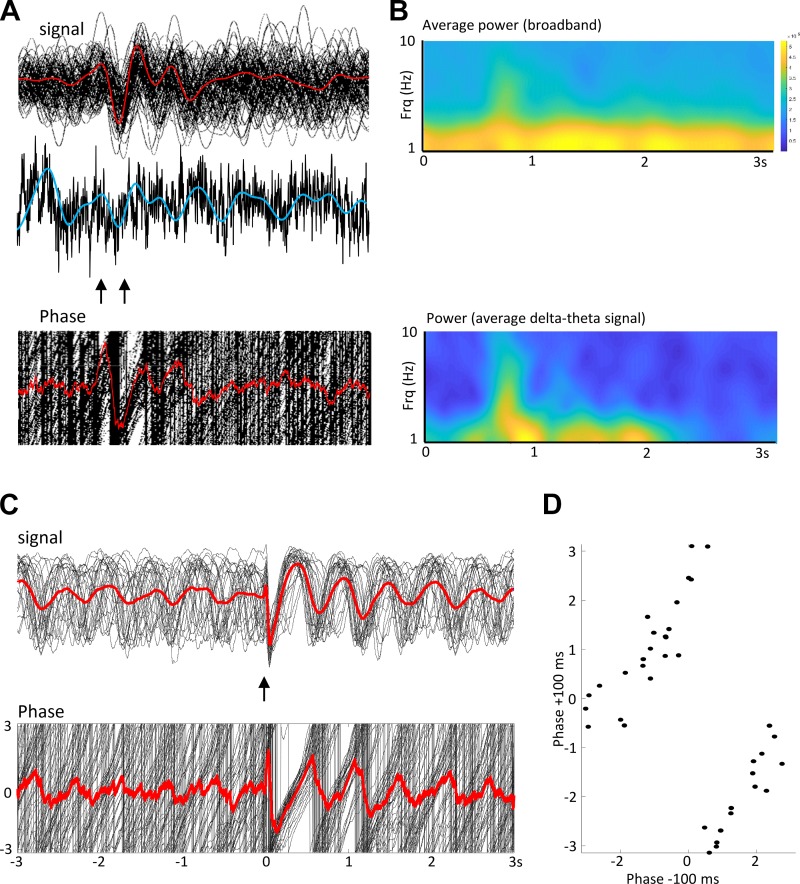
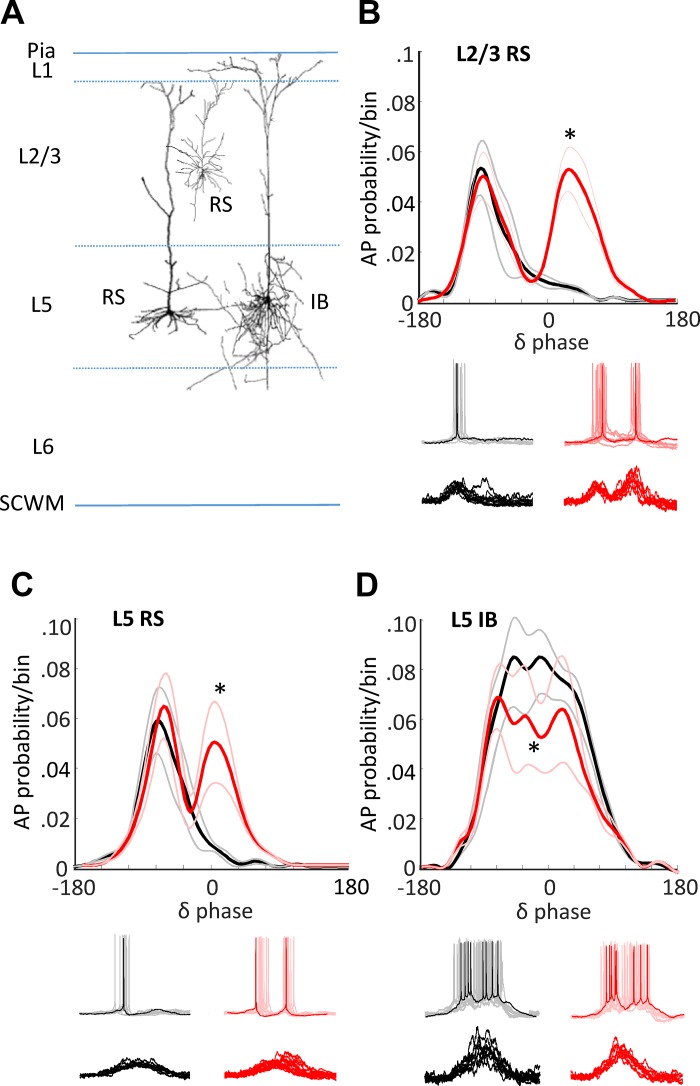
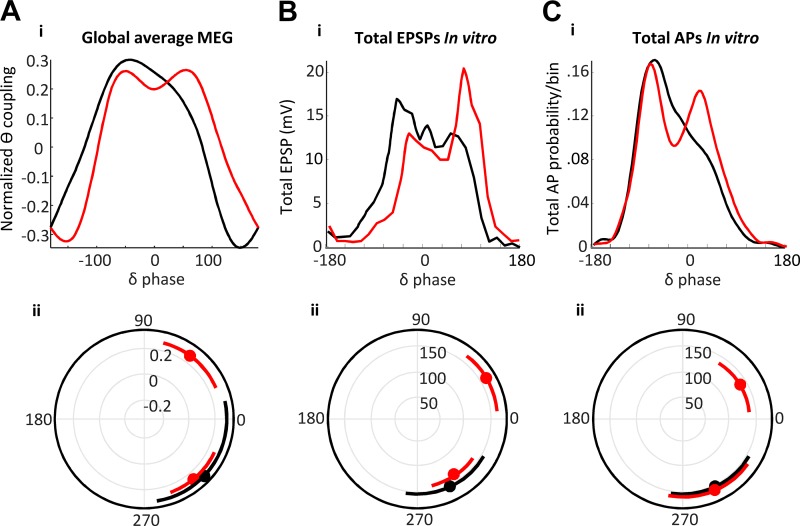
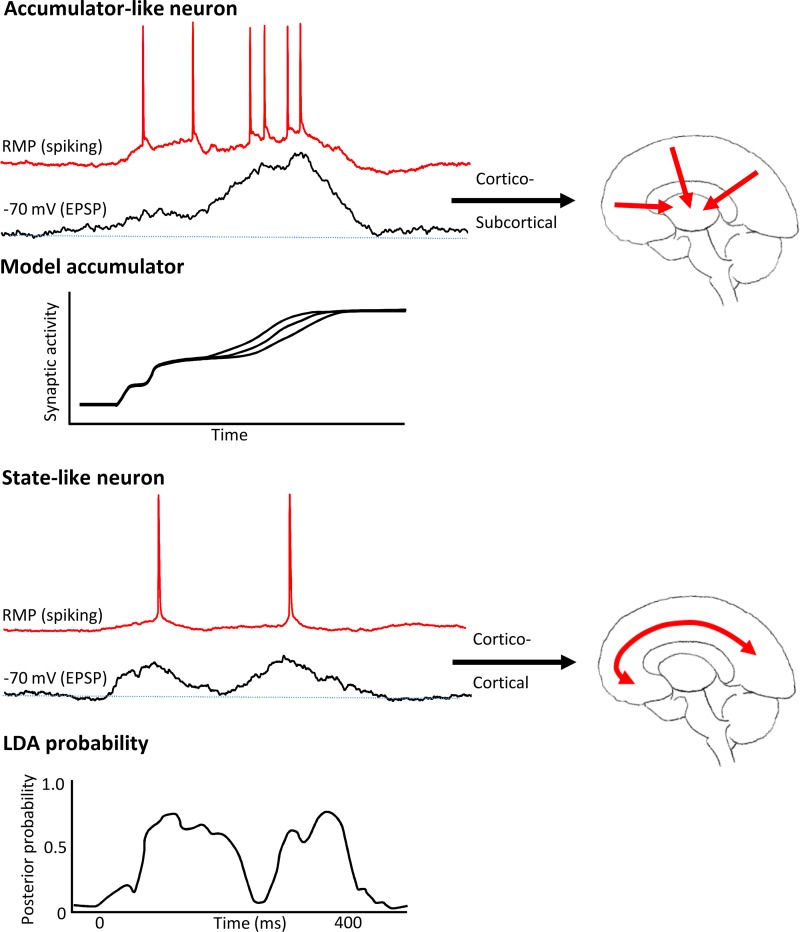
References
-
- Abeles M. Neural codes for higher brain function. In: Information Processing by the Brain, edited by Markowitsch HJ. Stuttgart, Germany: Hans Huber, 1988, p. 225–238.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources