

In silico Identification of Novel Toxin Homologs and Associated Mobile Genetic Elements in Clostridium perfringens
- PMID: 30699957
- PMCID: PMC6471305
- DOI: 10.3390/pathogens8010016
In silico Identification of Novel Toxin Homologs and Associated Mobile Genetic Elements in Clostridium perfringens
Abstract
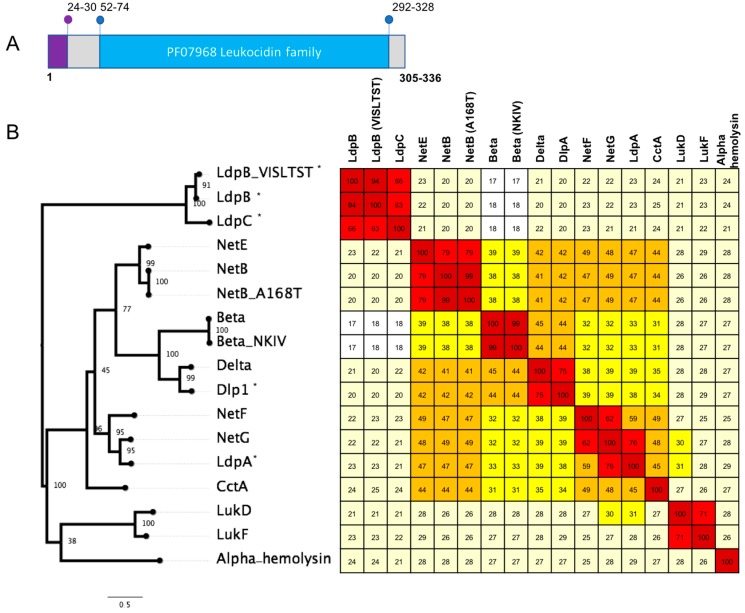
Clostridium perfringens causes a wide range of diseases in a variety of hosts, due to the production of a diverse set of toxins and extracellular enzymes. The C. perfringens toxins play an important role in pathogenesis, such that the presence and absence of the toxins is used as a typing scheme for the species. In recent years, several new toxins have been discovered that have been shown to be essential or highly correlated to diseases; these include binary enterotoxin (BecAB), NetB and NetF. In the current study, genome sequence analysis of C. perfringens isolates from diverse sources revealed several putative novel toxin homologs, some of which appeared to be associated with potential mobile genetic elements, including transposons and plasmids. Four novel toxin homologs encoding proteins related to the pore-forming Leukocidin/Hemolysin family were found in type A and G isolates. Two novel toxin homologs encoding proteins related to the epsilon aerolysin-like toxin family were identified in Type A and F isolates from humans, contaminated food and turkeys. A novel set of proteins related to clostridial binary toxins was also identified. While phenotypic characterisation is required before any of these homologs can be established as functional toxins, the in silico identification of these novel homologs on mobile genetic elements suggests the potential toxin reservoir of C. perfringens may be much larger than previously thought.
Keywords: Clostridium perfringens; binary toxin; epsilon; hemolysin; leukotoxin; pCP13; pCW3; plasmid; toxin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Analysis of the complete genome sequences of Clostridium perfringens strains harbouring the binary enterotoxin BEC gene and comparative genomics of pCP13-like family plasmids.BMC Genomics. 2022 Mar 23;23(1):226. doi: 10.1186/s12864-022-08453-4. BMC Genomics. 2022. PMID: 35321661 Free PMC article.
-
pCP13, a representative of a new family of conjugative toxin plasmids in Clostridium perfringens.Plasmid. 2019 Mar;102:37-45. doi: 10.1016/j.plasmid.2019.02.002. Epub 2019 Feb 18. Plasmid. 2019. PMID: 30790588
-
Genetic characteristics of toxigenic Clostridia and toxin gene evolution.Toxicon. 2013 Dec 1;75:63-89. doi: 10.1016/j.toxicon.2013.05.003. Epub 2013 May 23. Toxicon. 2013. PMID: 23707611
-
Pore-forming activity of clostridial binary toxins.Biochim Biophys Acta. 2016 Mar;1858(3):512-25. doi: 10.1016/j.bbamem.2015.08.006. Epub 2015 Aug 14. Biochim Biophys Acta. 2016. PMID: 26278641 Review.
-
Virulence Plasmids of Spore-Forming Bacteria.Microbiol Spectr. 2014 Dec;2(6):2.6.04. doi: 10.1128/microbiolspec.PLAS-0024-2014. Microbiol Spectr. 2014. PMID: 26104459 Free PMC article. Review.
Cited by
-
Genomic Diversity and Virulence Factors of Clostridium perfringens Isolated from Healthy and Necrotic Enteritis-Affected Broiler Chicken Farms in Quebec Province.Microorganisms. 2024 Dec 18;12(12):2624. doi: 10.3390/microorganisms12122624. Microorganisms. 2024. PMID: 39770825 Free PMC article.
-
Analysis of the complete genome sequences of Clostridium perfringens strains harbouring the binary enterotoxin BEC gene and comparative genomics of pCP13-like family plasmids.BMC Genomics. 2022 Mar 23;23(1):226. doi: 10.1186/s12864-022-08453-4. BMC Genomics. 2022. PMID: 35321661 Free PMC article.
-
Comparative in silico genome analysis of Clostridium perfringens unravels stable phylogroups with different genome characteristics and pathogenic potential.Sci Rep. 2021 Mar 24;11(1):6756. doi: 10.1038/s41598-021-86148-8. Sci Rep. 2021. PMID: 33762628 Free PMC article.
-
Cryo-EM structure of the octameric pore of Clostridium perfringens β-toxin.EMBO Rep. 2022 Dec 6;23(12):e54856. doi: 10.15252/embr.202254856. Epub 2022 Oct 10. EMBO Rep. 2022. PMID: 36215680 Free PMC article.
-
Necrotic enteritis and antibiotic-free production of broiler chickens: Challenges in testing and using alternative products.Anim Nutr. 2023 Dec 14;16:288-298. doi: 10.1016/j.aninu.2023.08.012. eCollection 2024 Mar. Anim Nutr. 2023. PMID: 38371475 Free PMC article. Review.
References
-
- Lacey J.A., Allnutt T.R., Vezina B., Van T.T.H., Stent T., Han X., Rood J.I., Wade B., Keyburn A.L., Seemann T., et al. Whole genome analysis reveals the diversity and evolutionary relationships between necrotic enteritis-causing strains of Clostridium perfringens. BMC Genom. 2018;19:379. doi: 10.1186/s12864-018-4771-1. - DOI - PMC - PubMed
-
- Lacey J.A., Keyburn A.L., Ford M.E., Portela R.W., Johanesen P.A., Lyras D., Moore R.J. Conjugation-mediated horizontal gene transfer of Clostridium perfringens plasmids in the chicken gastrointestinal tract results in the formation of new virulent strains. Appl. Environ. Microbiol. 2017;83:e01814-17. doi: 10.1128/AEM.01814-17. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
