

Deleting Full Length Titin Versus the Titin M-Band Region Leads to Differential Mechanosignaling and Cardiac Phenotypes
- PMID: 30700140
- PMCID: PMC6453709
- DOI: 10.1161/CIRCULATIONAHA.118.037588
Deleting Full Length Titin Versus the Titin M-Band Region Leads to Differential Mechanosignaling and Cardiac Phenotypes
Abstract
Background: Titin is a giant elastic protein that spans the half-sarcomere from Z-disk to M-band. It acts as a molecular spring and mechanosensor and has been linked to striated muscle disease. The pathways that govern titin-dependent cardiac growth and contribute to disease are diverse and difficult to dissect.
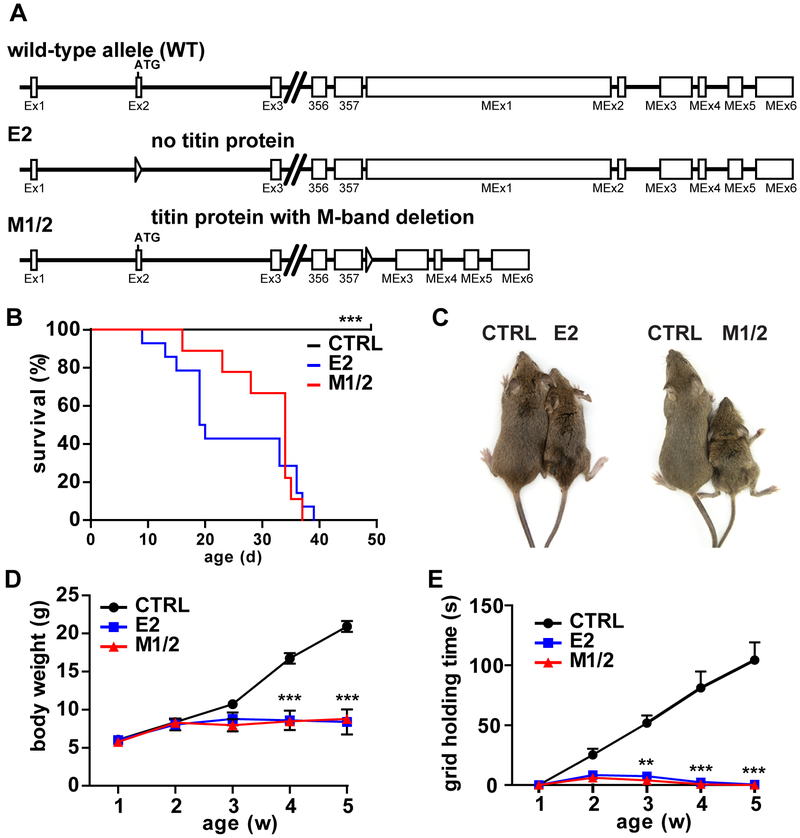
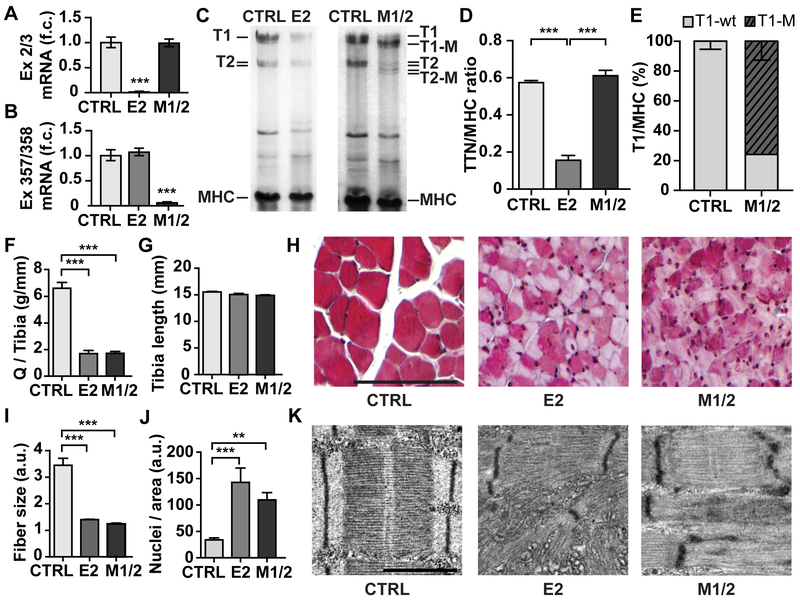
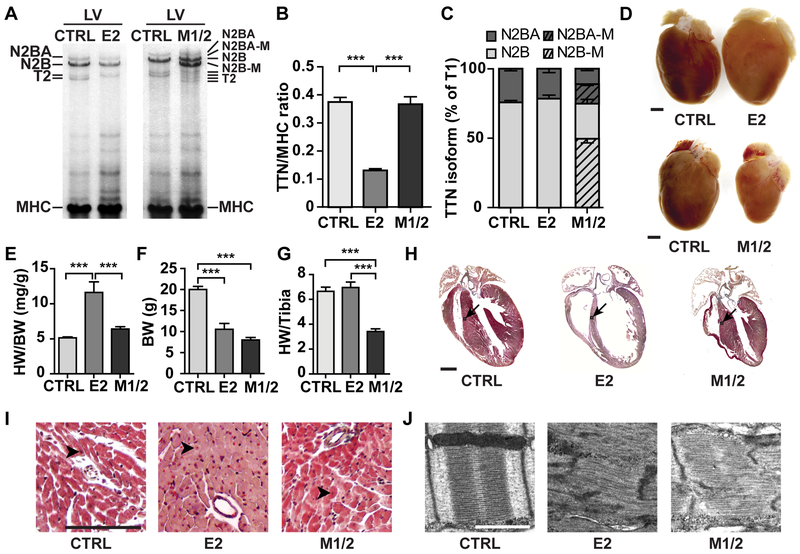
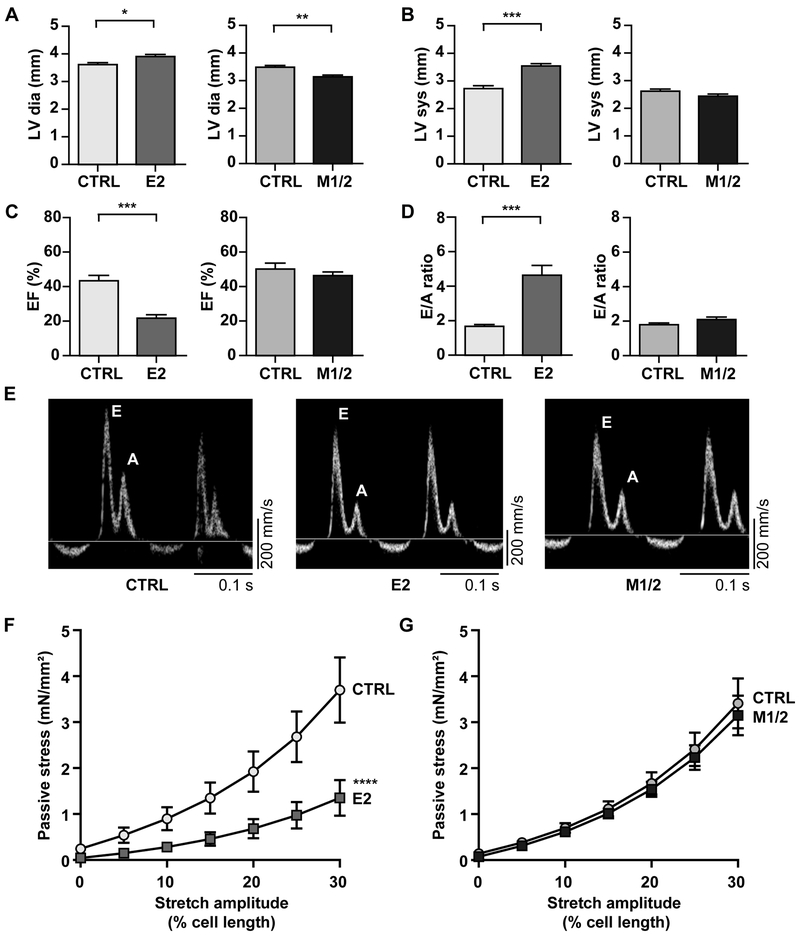
Methods: To study titin deficiency versus dysfunction, the authors generated and compared striated muscle specific knockouts (KOs) with progressive postnatal loss of the complete titin protein by removing exon 2 (E2-KO) or an M-band truncation that eliminates proper sarcomeric integration, but retains all other functional domains (M-band exon 1/2 [M1/2]-KO). The authors evaluated cardiac function, cardiomyocyte mechanics, and the molecular basis of the phenotype.
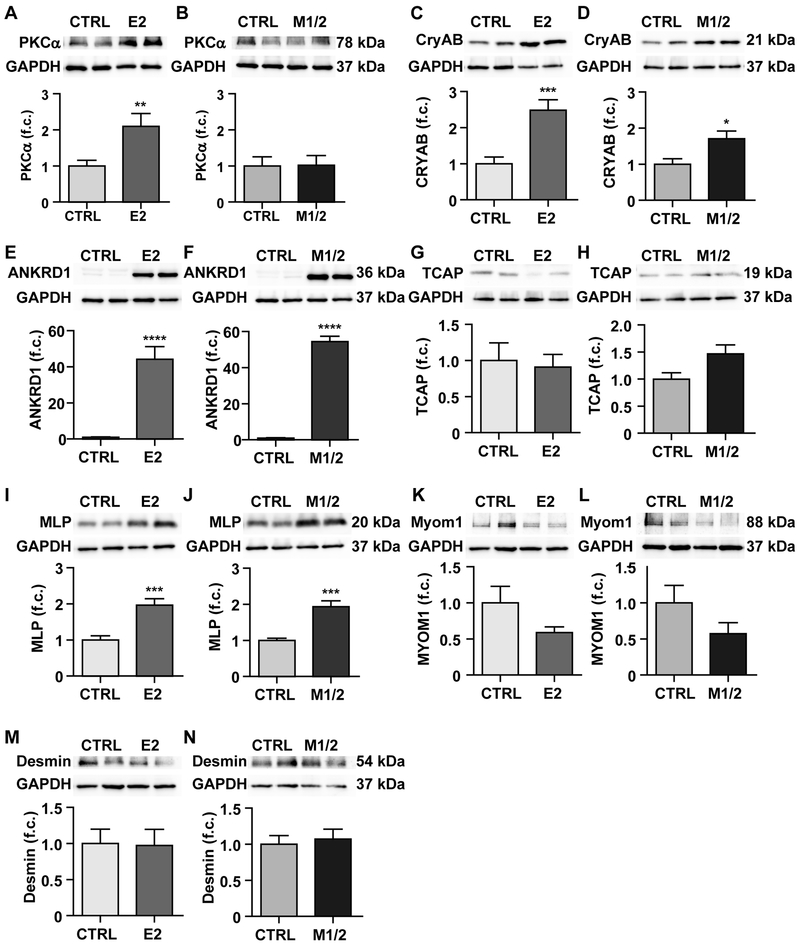
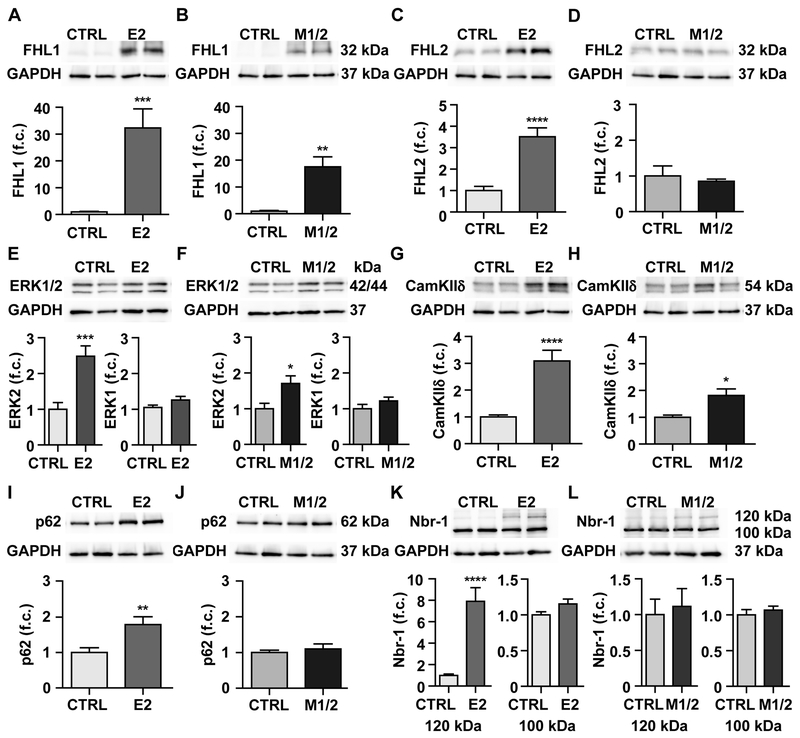
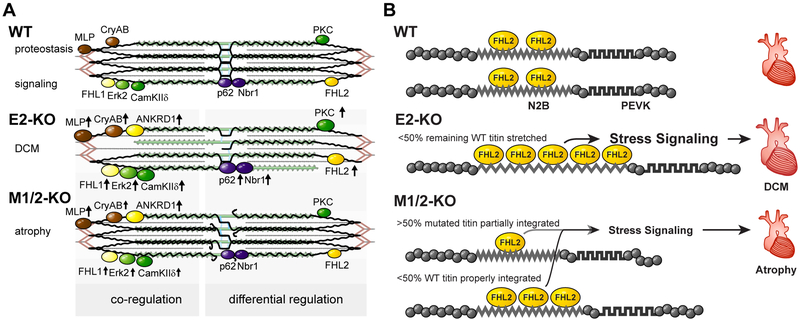
Results: Skeletal muscle atrophy with reduced strength, severe sarcomere disassembly, and lethality from 2 weeks of age were shared between the models. Cardiac phenotypes differed considerably: loss of titin leads to dilated cardiomyopathy with combined systolic and diastolic dysfunction-the absence of M-band titin to cardiac atrophy and preserved function. The elastic properties of M1/2-KO cardiomyocytes are maintained, while passive stiffness is reduced in the E2-KO. In both KOs, we find an increased stress response and increased expression of proteins linked to titin-based mechanotransduction (CryAB, ANKRD1, muscle LIM protein, FHLs, p42, Camk2d, p62, and Nbr1). Among them, FHL2 and the M-band signaling proteins p62 and Nbr1 are exclusively upregulated in the E2-KO, suggesting a role in the differential pathology of titin truncation versus deficiency of the full-length protein. The differential stress response is consistent with truncated titin contributing to the mechanical properties in M1/2-KOs, while low titin levels in E2-KOs lead to reduced titin-based stiffness and increased strain on the remaining titin molecules.
Conclusions: Progressive depletion of titin leads to sarcomere disassembly and atrophy in striated muscle. In the complete knockout, remaining titin molecules experience increased strain, resulting in mechanically induced trophic signaling and eventually dilated cardiomyopathy. The truncated titin in M1/2-KO helps maintain the passive properties and thus reduces mechanically induced signaling. Together, these findings contribute to the molecular understanding of why titin mutations differentially affect cardiac growth and have implications for genotype-phenotype relations that support a personalized medicine approach to the diverse titinopathies.
Keywords: cardiomyopathies; heart diseases; hypertrophy; models, animal; muscles; myocardial contraction.
Figures
Similar articles
-
Targeted deletion of titin N2B region leads to diastolic dysfunction and cardiac atrophy.Proc Natl Acad Sci U S A. 2007 Feb 27;104(9):3444-9. doi: 10.1073/pnas.0608543104. Epub 2007 Feb 20. Proc Natl Acad Sci U S A. 2007. PMID: 17360664 Free PMC article.
-
Deleting titin's I-band/A-band junction reveals critical roles for titin in biomechanical sensing and cardiac function.Proc Natl Acad Sci U S A. 2014 Oct 7;111(40):14589-94. doi: 10.1073/pnas.1411493111. Epub 2014 Sep 22. Proc Natl Acad Sci U S A. 2014. PMID: 25246556 Free PMC article.
-
Sense and stretchability: the role of titin and titin-associated proteins in myocardial stress-sensing and mechanical dysfunction.Cardiovasc Res. 2008 Mar 1;77(4):637-48. doi: 10.1016/j.cardiores.2007.03.029. Cardiovasc Res. 2008. PMID: 17475230 Review.
-
Muscle atrophy in titin M-line deficient mice.J Muscle Res Cell Motil. 2005;26(6-8):381-8. doi: 10.1007/s10974-005-9020-y. J Muscle Res Cell Motil. 2005. PMID: 16470336
-
Titin Gene and Protein Functions in Passive and Active Muscle.Annu Rev Physiol. 2018 Feb 10;80:389-411. doi: 10.1146/annurev-physiol-021317-121234. Epub 2017 Nov 13. Annu Rev Physiol. 2018. PMID: 29131758 Review.
Cited by
-
PIK3R1, SPNB2, and CRYAB as Potential Biomarkers for Patients with Diabetes and Developing Acute Myocardial Infarction.Int J Endocrinol. 2021 Nov 30;2021:2267736. doi: 10.1155/2021/2267736. eCollection 2021. Int J Endocrinol. 2021. PMID: 34887920 Free PMC article.
-
Titin mutations and muscle disease.Pflugers Arch. 2019 May;471(5):673-682. doi: 10.1007/s00424-019-02272-5. Epub 2019 Mar 27. Pflugers Arch. 2019. PMID: 30919088 Free PMC article. Review.
-
Mechanically knocking out titin reveals protein tension loss as a trigger of muscle disease.Nat Biomed Eng. 2025 Jun 5. doi: 10.1038/s41551-025-01403-x. Online ahead of print. Nat Biomed Eng. 2025. PMID: 40473933
-
Does Myocardial Atrophy Represent Anti-Arrhythmic Phenotype?Biomedicines. 2022 Nov 4;10(11):2819. doi: 10.3390/biomedicines10112819. Biomedicines. 2022. PMID: 36359339 Free PMC article. Review.
-
Cardiomyocyte Dysfunction in Inherited Cardiomyopathies.Int J Mol Sci. 2021 Oct 15;22(20):11154. doi: 10.3390/ijms222011154. Int J Mol Sci. 2021. PMID: 34681814 Free PMC article. Review.
References
-
- Herman DS, Lam L, Taylor MRG, Wang L, Teekakirikul P, Christodoulou D, Conner L, DePalma SR, McDonough B, Sparks E, Teodorescu DL, Cirino AL, Banner NR, Pennell DJ, Graw S, Merlo M, Di Lenarda A, Sinagra G, Bos JM, Ackerman MJ, Mitchell RN, Murry CE, Lakdawala NK, Ho CY, Barton PJR, Cook SA, Mestroni L, Seidman JG, Seidman CE. Truncations of titin causing dilated cardiomyopathy. N Engl J Med. 2012;366:619–628. - PMC - PubMed
-
- Hackman P, Marchand S, Sarparanta J, Vihola A, Pénisson-Besnier I, Eymard B, Pardal-Fernández JM, Hammouda E-H, Richard I, Illa I, Udd B. Truncating mutations in C-terminal titin may cause more severe tibial muscular dystrophy (TMD). Neuromuscul Disord NMD. 2008;18:922–928. - PubMed
-
- Mues A, van der Ven PF, Young P, Fürst DO, Gautel M. Two immunoglobulin-like domains of the Z-disc portion of titin interact in a conformation-dependent way with telethonin. FEBS Lett. 1998;428:111–114. - PubMed
-
- Labeit S, Kolmerer B. Titins: giant proteins in charge of muscle ultrastructure and elasticity. Science. 1995;270:293–296. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
