

Microtubule stabilizer reveals requirement of Ca2+-dependent conformational changes of microtubules for rapid coiling of haptonema in haptophyte algae
- PMID: 30700402
- PMCID: PMC6398456
- DOI: 10.1242/bio.036590
Microtubule stabilizer reveals requirement of Ca2+-dependent conformational changes of microtubules for rapid coiling of haptonema in haptophyte algae
Abstract
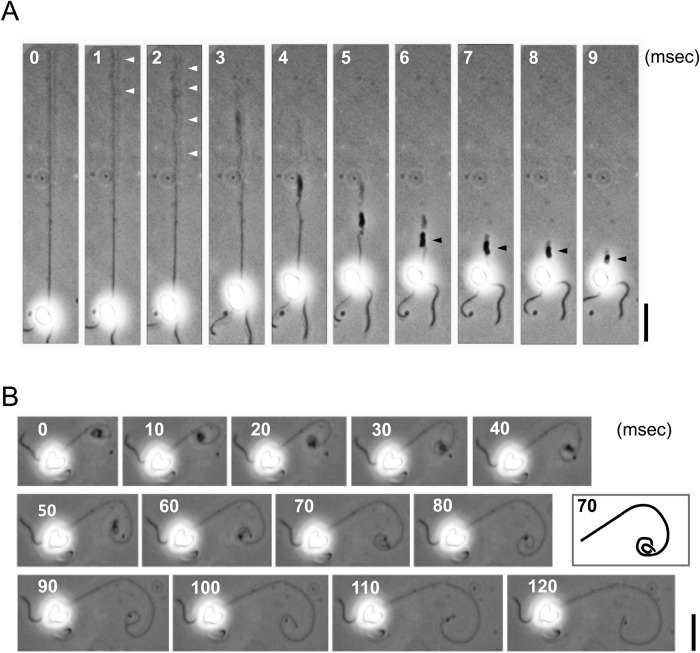
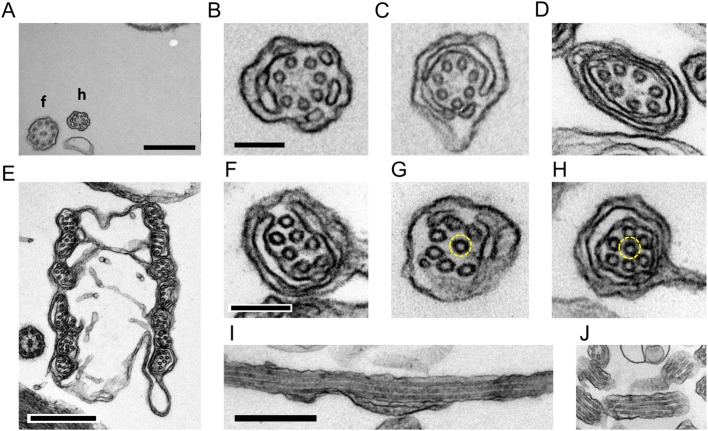
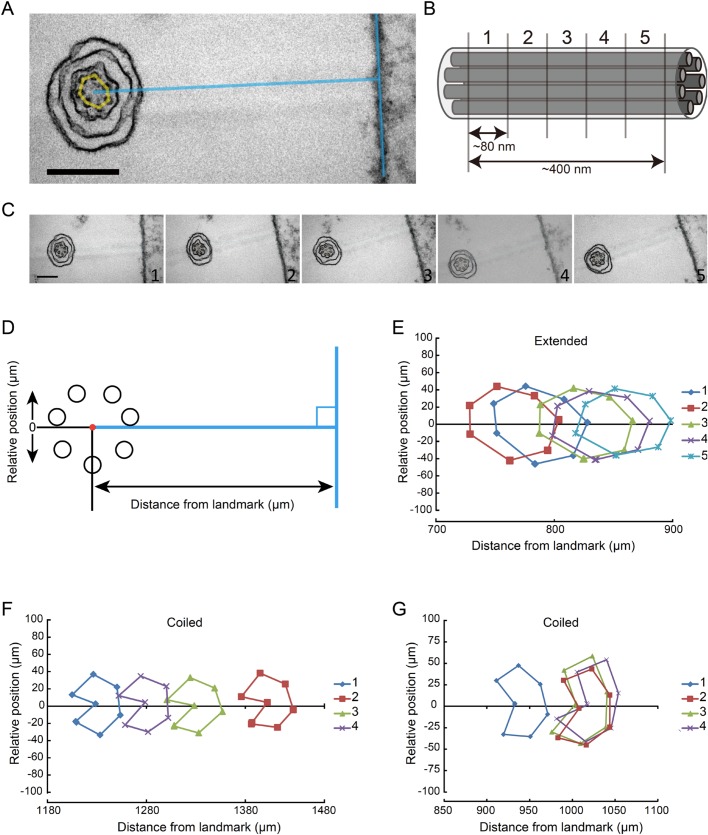
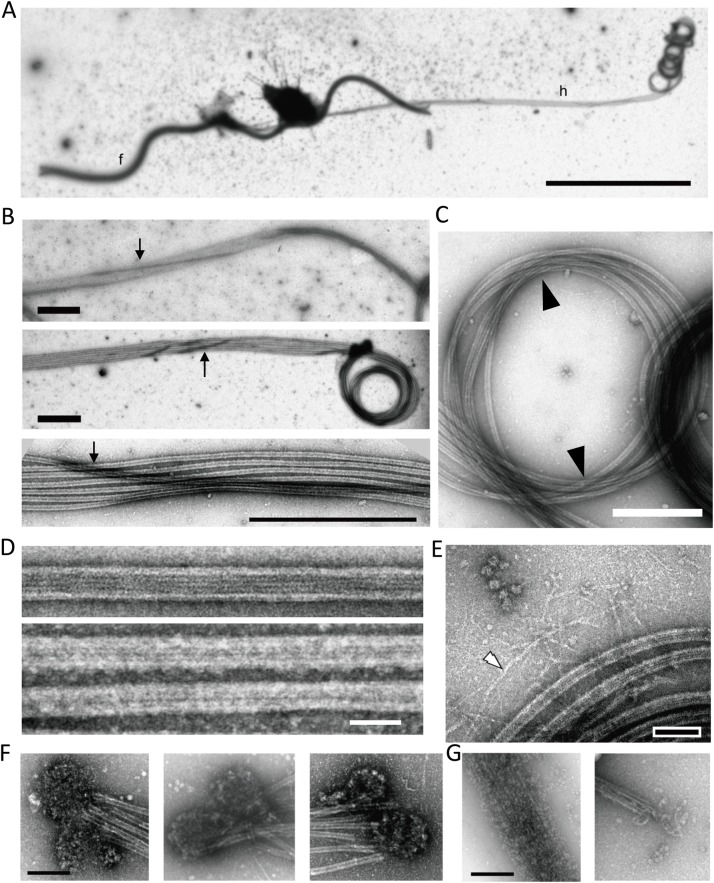
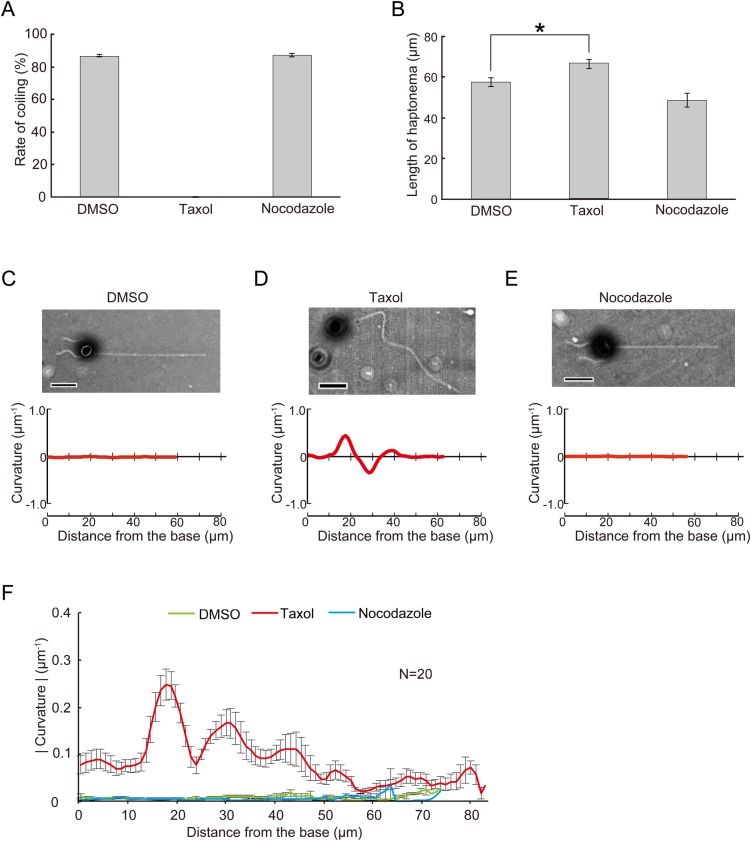
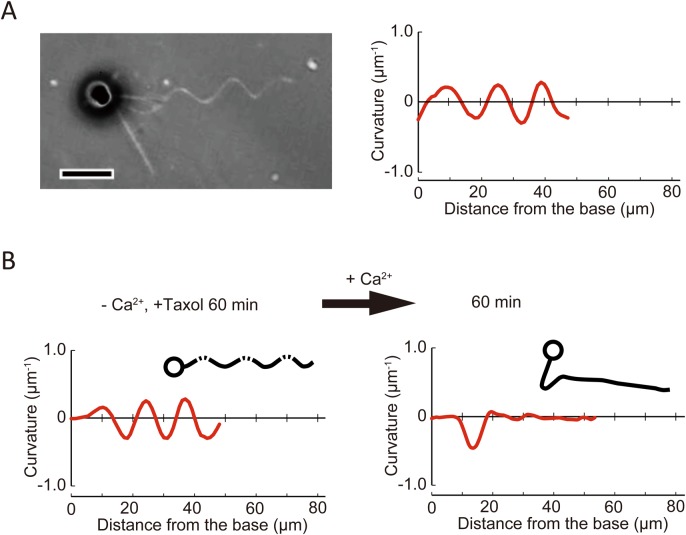
A haptonema is an elongated microtubule-based motile organelle uniquely present in haptophytes. The most notable and rapid movement of a haptonema is 'coiling', which occurs within a few milliseconds following mechanical stimulation in an unknown motor-independent mechanism. Here, we analyzed the coiling process in detail by high-speed filming and showed that haptonema coiling was initiated by left-handed twisting of the haptonema, followed by writhing to form a helix from the distal tip. On recovery from a mechanical stimulus, the helix slowly uncoiled from the proximal region. Electron microscopy showed that the seven microtubules in a haptonema were arranged mostly in parallel but that one of the microtubules often wound around the others in the extended state. A microtubule stabilizer, paclitaxel, inhibited coiling and induced right-handed twisting of the haptonema in the absence of Ca2+, suggesting changes in the mechanical properties of microtubules. Addition of Ca2+ resulted in the conversion of haptonematal twist into the planar bends near the proximal region. These results indicate that switching microtubule conformation, possibly with the aid of Ca2+-binding microtubule-associated proteins is responsible for rapid haptonematal coiling.
Keywords: Chrysochromulina; Curvature; Haptonema; Haptophyte; Microtubule; Paclitaxel.
© 2019. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
Similar articles
-
Taxol causes rapid gross structural rearrangement of a native microtubule bundle.Cell Biochem Funct. 1994 Sep;12(3):191-200. doi: 10.1002/cbf.290120307. Cell Biochem Funct. 1994. PMID: 7955129
-
A haptophyte bearing siliceous scales: ultrastructure and phylogenetic position of Hyalolithus neolepis gen. et sp. nov. (Prymnesiophyceae, Haptophyta).Protist. 2006 Jun;157(2):213-34. doi: 10.1016/j.protis.2006.02.004. Epub 2006 May 2. Protist. 2006. PMID: 16647294
-
Mechanical effects of EB1 on microtubules depend on GTP hydrolysis state and presence of paclitaxel.Cytoskeleton (Hoboken). 2014 Sep;71(9):530-41. doi: 10.1002/cm.21190. Epub 2014 Sep 12. Cytoskeleton (Hoboken). 2014. PMID: 25160006
-
Lis1 restricts the conformational changes in cytoplasmic dynein on microtubules.Microscopy (Oxf). 2015 Dec;64(6):419-27. doi: 10.1093/jmicro/dfv055. Epub 2015 Sep 14. Microscopy (Oxf). 2015. PMID: 26371280
-
Ca/Ba/Sr-induced conformational changes of ciliary axonemes.Cell Motil Cytoskeleton. 1990;17(3):187-96. doi: 10.1002/cm.970170306. Cell Motil Cytoskeleton. 1990. PMID: 1980094
Cited by
-
A Novel Single-Domain Na+-Selective Voltage-Gated Channel in Photosynthetic Eukaryotes.Plant Physiol. 2020 Dec;184(4):1674-1683. doi: 10.1104/pp.20.00889. Epub 2020 Oct 1. Plant Physiol. 2020. PMID: 33004614 Free PMC article.
-
Tree of motility - A proposed history of motility systems in the tree of life.Genes Cells. 2020 Jan;25(1):6-21. doi: 10.1111/gtc.12737. Genes Cells. 2020. PMID: 31957229 Free PMC article. Review.
-
Origins of eukaryotic excitability.Philos Trans R Soc Lond B Biol Sci. 2021 Mar 15;376(1820):20190758. doi: 10.1098/rstb.2019.0758. Epub 2021 Jan 25. Philos Trans R Soc Lond B Biol Sci. 2021. PMID: 33487111 Free PMC article. Review.
References
-
- Amos W. B., Routledge L. M. and Yew F. F. (1975). Calcium-binding proteins in a Vorticellid contractile organelle. J. Cell Sci. 19, 203-213. - PubMed
-
- Anderson R. A. and Kawachi M. (2005). Traditional microalgae isolation techniques. In Algal Culturing Techniques (ed. Anderson R. A.), pp. 83-100. Amsterdam: Elsevier Academic Press.
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
