

Necessity and Contingency in Developmental Genetic Screens: EGF, Wnt, and Semaphorin Pathways in Vulval Induction of the Nematode Oscheius tipulae
- PMID: 30700527
- PMCID: PMC6456316
- DOI: 10.1534/genetics.119.301970
Necessity and Contingency in Developmental Genetic Screens: EGF, Wnt, and Semaphorin Pathways in Vulval Induction of the Nematode Oscheius tipulae
Abstract
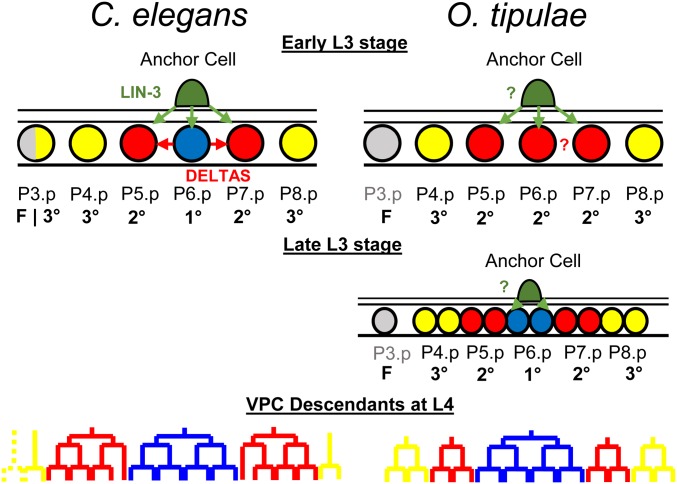
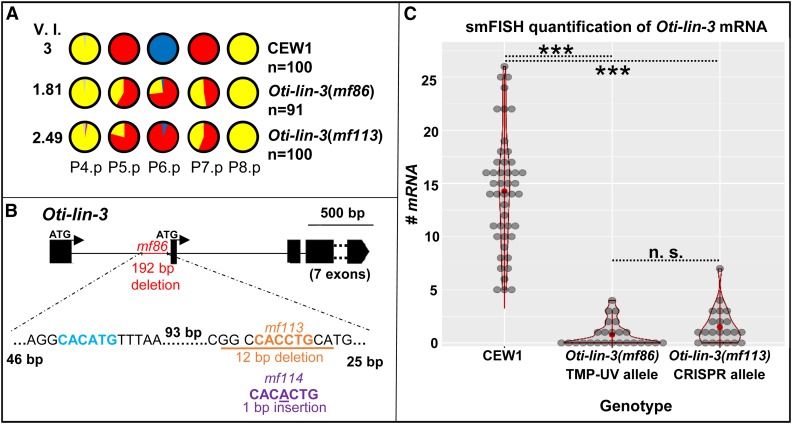
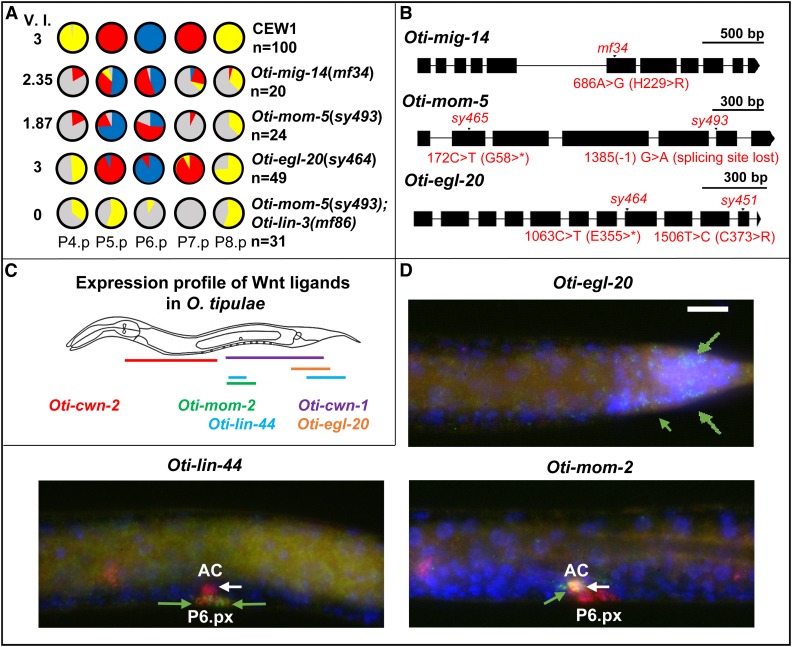
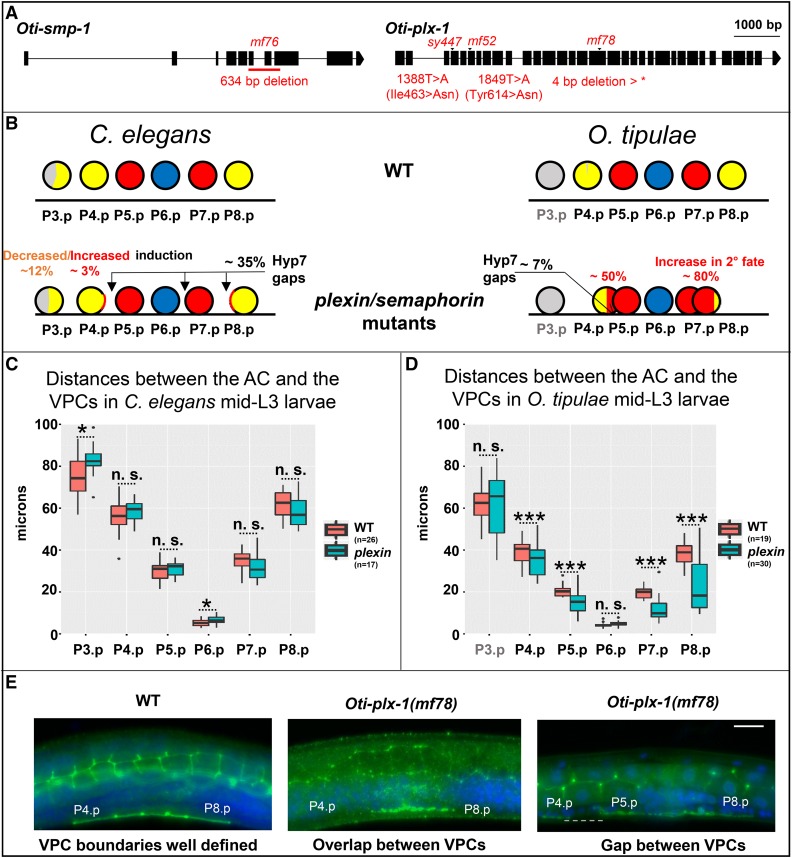
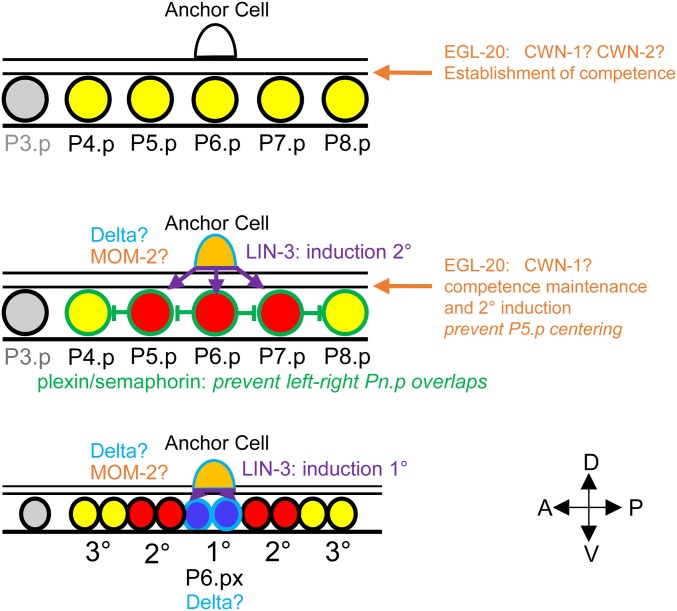
Genetic screens in the nematode Caenorhabditis elegans identified the EGF/Ras and Notch pathways as central for vulval precursor cell fate patterning. Schematically, the anchor cell secretes EGF, inducing the P6.p cell to a primary (1°) vulval fate; P6.p in turn induces its neighbors to a secondary (2°) fate through Delta-Notch signaling and represses Ras signaling. In the nematode Oscheius tipulae, the anchor cell successively induces 2° then 1° vulval fates. Here, we report on the molecular identification of mutations affecting vulval induction in O. tipulae A single Induction Vulvaless mutation was found, which we identify as a cis-regulatory deletion in a tissue-specific enhancer of the O. tipulae lin-3 homolog, confirmed by clustered regularly interspaced short palindromic repeats/Cas9 mutation. In contrast to this predictable Vulvaless mutation, mutations resulting in an excess of 2° fates unexpectedly correspond to the plexin/semaphorin pathway. Hyperinduction of P4.p and P8.p in these mutants likely results from mispositioning of these cells due to a lack of contact inhibition. The third signaling pathway found by forward genetics in O. tipulae is the Wnt pathway; a decrease in Wnt pathway activity results in loss of vulval precursor competence and induction, and 1° fate miscentering on P5.p. Our results suggest that the EGF and Wnt pathways have qualitatively similar activities in vulval induction in C. elegans and O. tipulae, albeit with quantitative differences in the effects of mutation. Thus, the derived induction process in C. elegans with an early induction of the 1° fate appeared during evolution, after the recruitment of the EGF pathway for vulval induction.
Keywords: C. elegans; O. tipulae; Wnt; evolution of development; genetic screens; lin-3/EGF; plexin; semaphorin.
Copyright © 2019 Vargas-Velazquez et al.
Figures
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
