

Pluripotency and X chromosome dynamics revealed in pig pre-gastrulating embryos by single cell analysis
- PMID: 30700715
- PMCID: PMC6353908
- DOI: 10.1038/s41467-019-08387-8
Pluripotency and X chromosome dynamics revealed in pig pre-gastrulating embryos by single cell analysis
Abstract
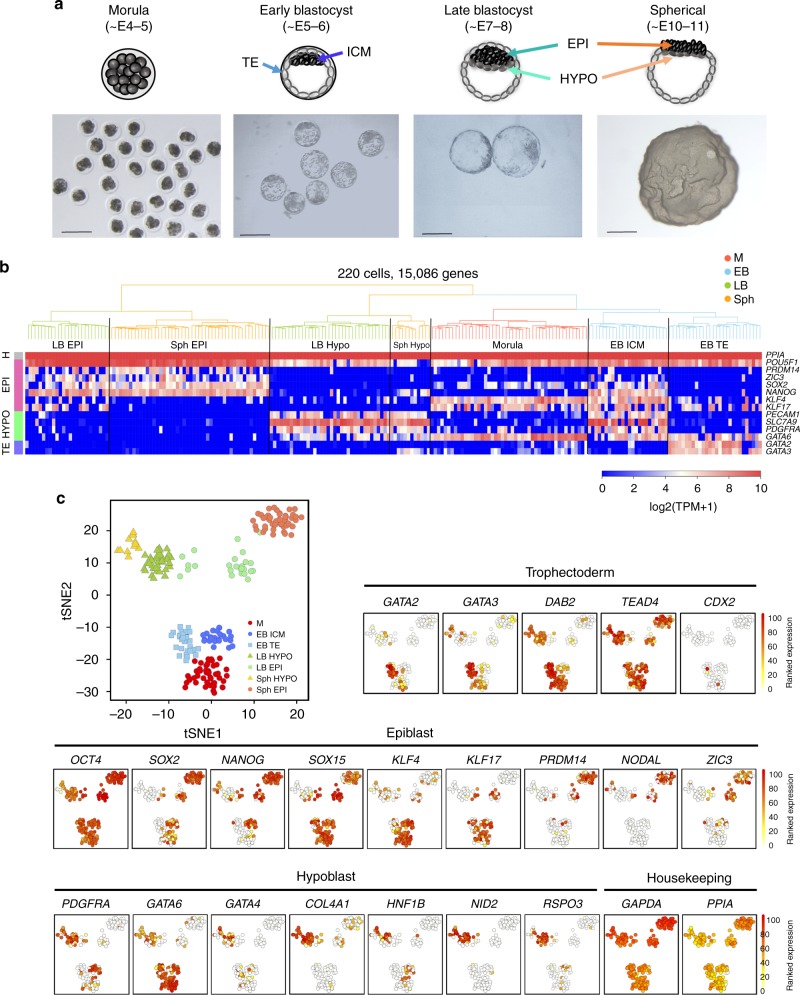
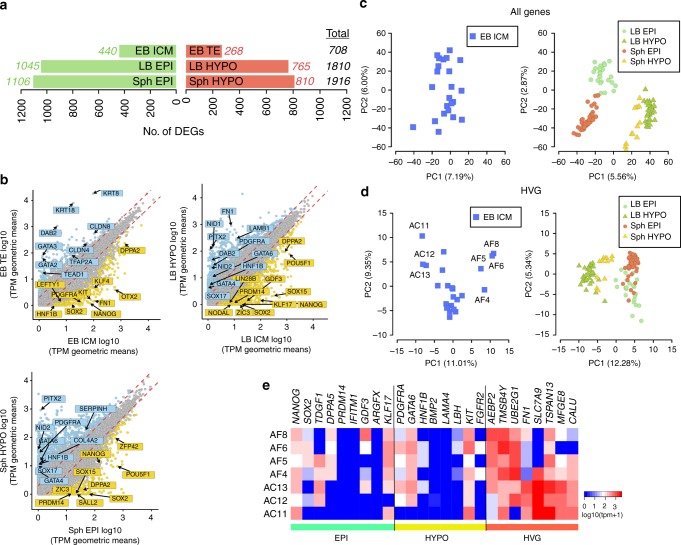
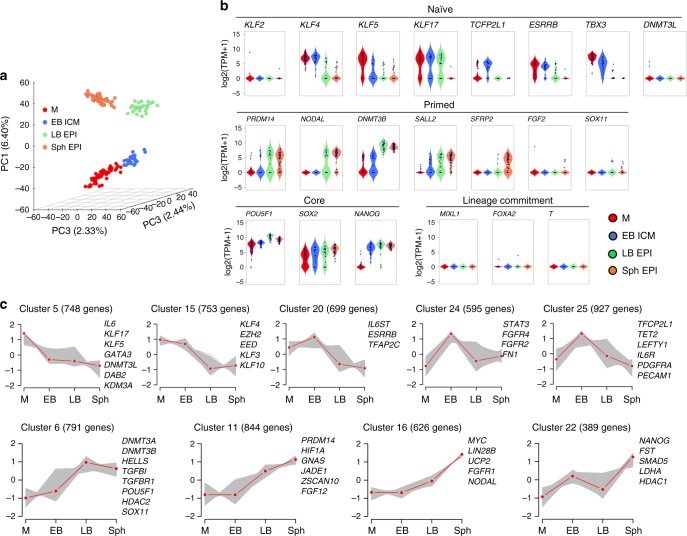
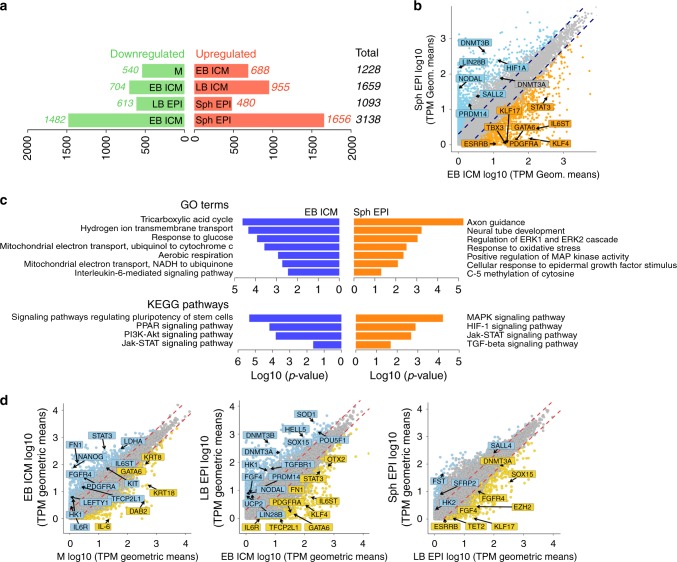
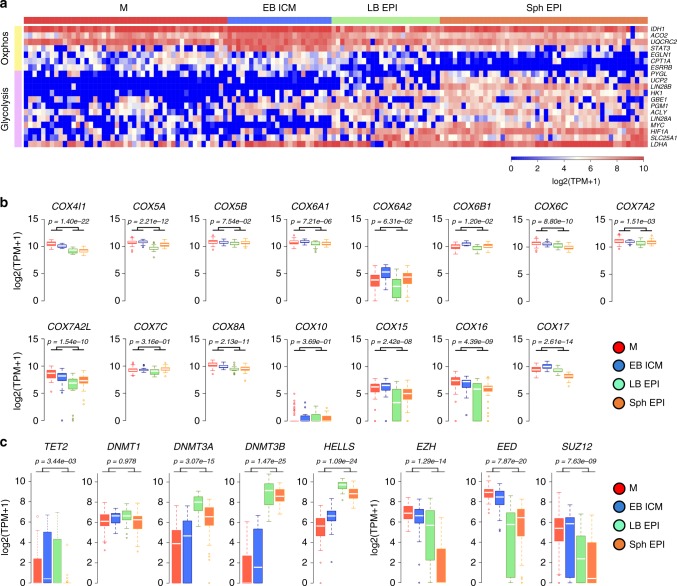
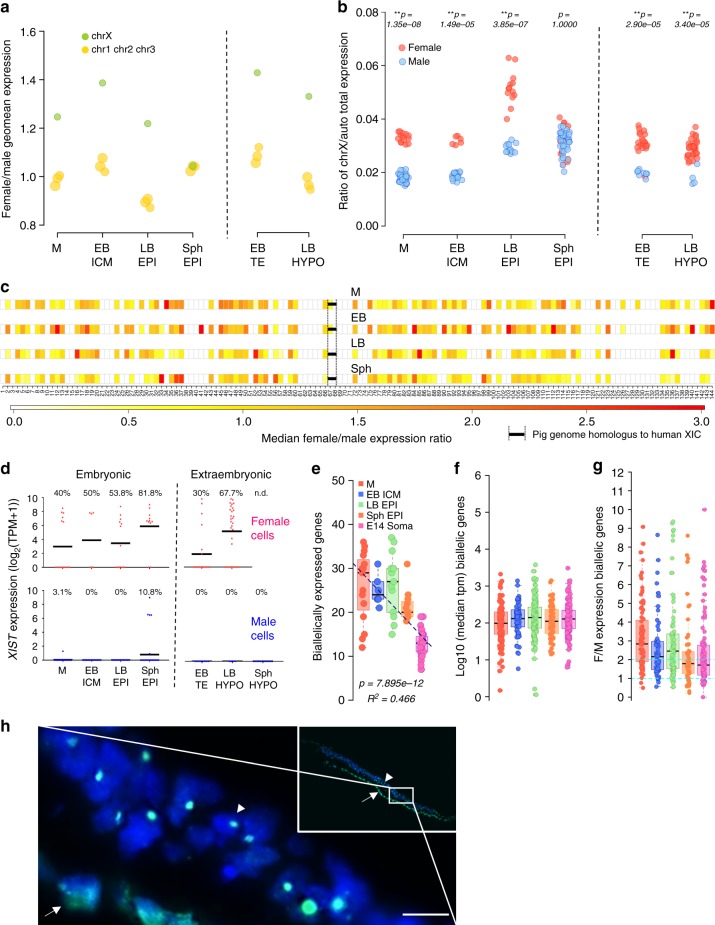
High-resolution molecular programmes delineating the cellular foundations of mammalian embryogenesis have emerged recently. Similar analysis of human embryos is limited to pre-implantation stages, since early post-implantation embryos are largely inaccessible. Notwithstanding, we previously suggested conserved principles of pig and human early development. For further insight on pluripotent states and lineage delineation, we analysed pig embryos at single cell resolution. Here we show progressive segregation of inner cell mass and trophectoderm in early blastocysts, and of epiblast and hypoblast in late blastocysts. We show that following an emergent short naive pluripotent signature in early embryos, there is a protracted appearance of a primed signature in advanced embryonic stages. Dosage compensation with respect to the X-chromosome in females is attained via X-inactivation in late epiblasts. Detailed human-pig comparison is a basis towards comprehending early human development and a foundation for further studies of human pluripotent stem cell differentiation in pig interspecies chimeras.
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Lineage specification and pluripotency revealed by transcriptome analysis from oocyte to blastocyst in pig.FASEB J. 2020 Jan;34(1):691-705. doi: 10.1096/fj.201901818RR. Epub 2019 Nov 26. FASEB J. 2020. PMID: 31914626
-
Live imaging of X chromosome reactivation dynamics in early mouse development can discriminate naïve from primed pluripotent stem cells.Development. 2016 Aug 15;143(16):2958-64. doi: 10.1242/dev.136739. Epub 2016 Jul 28. Development. 2016. PMID: 27471261
-
A single-cell transcriptome atlas of marsupial embryogenesis and X inactivation.Nature. 2020 Oct;586(7830):612-617. doi: 10.1038/s41586-020-2629-6. Epub 2020 Aug 19. Nature. 2020. PMID: 32814901 Free PMC article.
-
Epiblast stem cells contribute new insight into pluripotency and gastrulation.Dev Growth Differ. 2010 Apr;52(3):293-301. doi: 10.1111/j.1440-169X.2010.01171.x. Epub 2010 Mar 7. Dev Growth Differ. 2010. PMID: 20298258 Review.
-
Stepwise pluripotency transitions in mouse stem cells.EMBO Rep. 2022 Sep 5;23(9):e55010. doi: 10.15252/embr.202255010. Epub 2022 Jul 29. EMBO Rep. 2022. PMID: 35903955 Free PMC article. Review.
Cited by
-
Contiguous erosion of the inactive X in human pluripotency concludes with global DNA hypomethylation.Cell Rep. 2021 Jun 8;35(10):109215. doi: 10.1016/j.celrep.2021.109215. Cell Rep. 2021. PMID: 34107261 Free PMC article.
-
Intestinal single-cell atlas reveals novel lymphocytes in pigs with similarities to human cells.Life Sci Alliance. 2022 Aug 22;5(10):e202201442. doi: 10.26508/lsa.202201442. Print 2022 Oct. Life Sci Alliance. 2022. PMID: 35995567 Free PMC article.
-
Generation and characterization of stable pig pregastrulation epiblast stem cell lines.Cell Res. 2022 Apr;32(4):383-400. doi: 10.1038/s41422-021-00592-9. Epub 2021 Nov 30. Cell Res. 2022. PMID: 34848870 Free PMC article.
-
No imprinted XIST expression in pigs: biallelic XIST expression in early embryos and random X inactivation in placentas.Cell Mol Life Sci. 2019 Nov;76(22):4525-4538. doi: 10.1007/s00018-019-03123-3. Epub 2019 May 28. Cell Mol Life Sci. 2019. PMID: 31139846 Free PMC article.
-
Establishment of porcine embryonic stem cells in simplified serum free media and feeder free expansion.Stem Cell Res Ther. 2024 Aug 7;15(1):245. doi: 10.1186/s13287-024-03858-2. Stem Cell Res Ther. 2024. PMID: 39113095 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
