

Template-directed RNA polymerization and enhanced ribozyme catalysis inside membraneless compartments formed by coacervates
- PMID: 30700721
- PMCID: PMC6353945
- DOI: 10.1038/s41467-019-08353-4
Template-directed RNA polymerization and enhanced ribozyme catalysis inside membraneless compartments formed by coacervates
Abstract
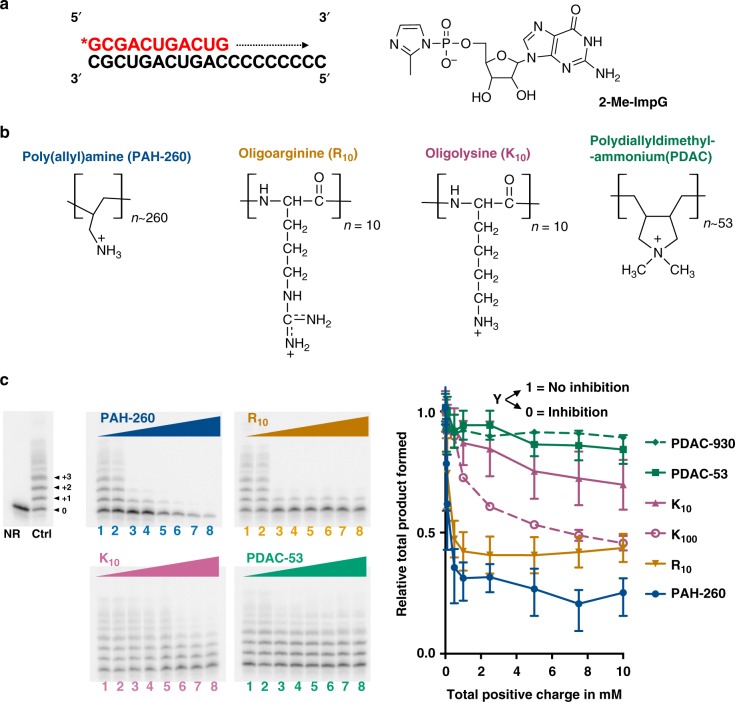
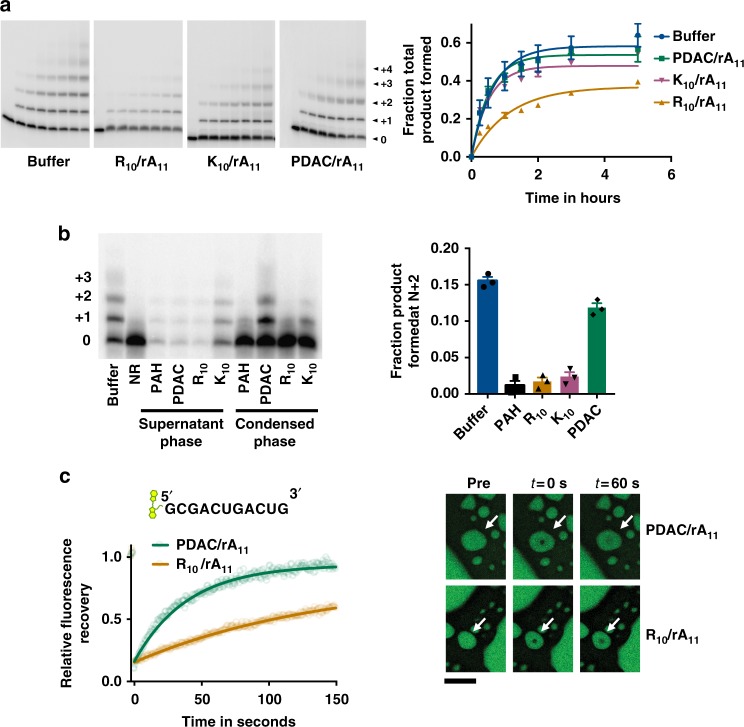
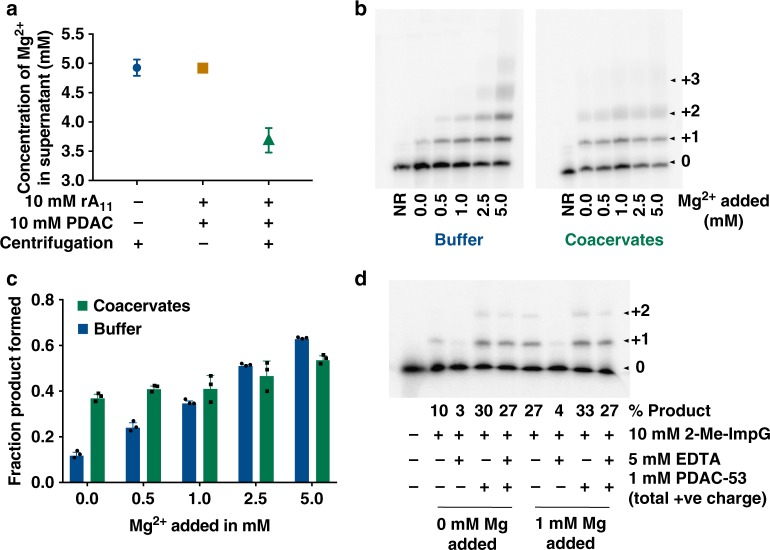
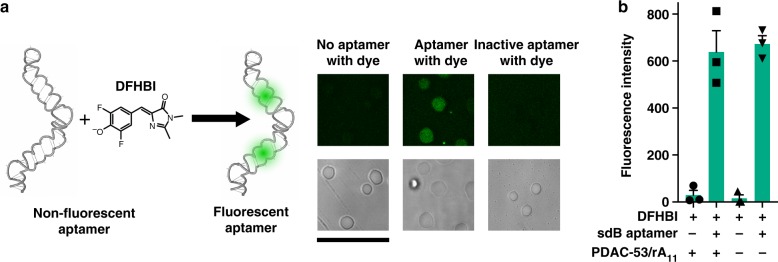
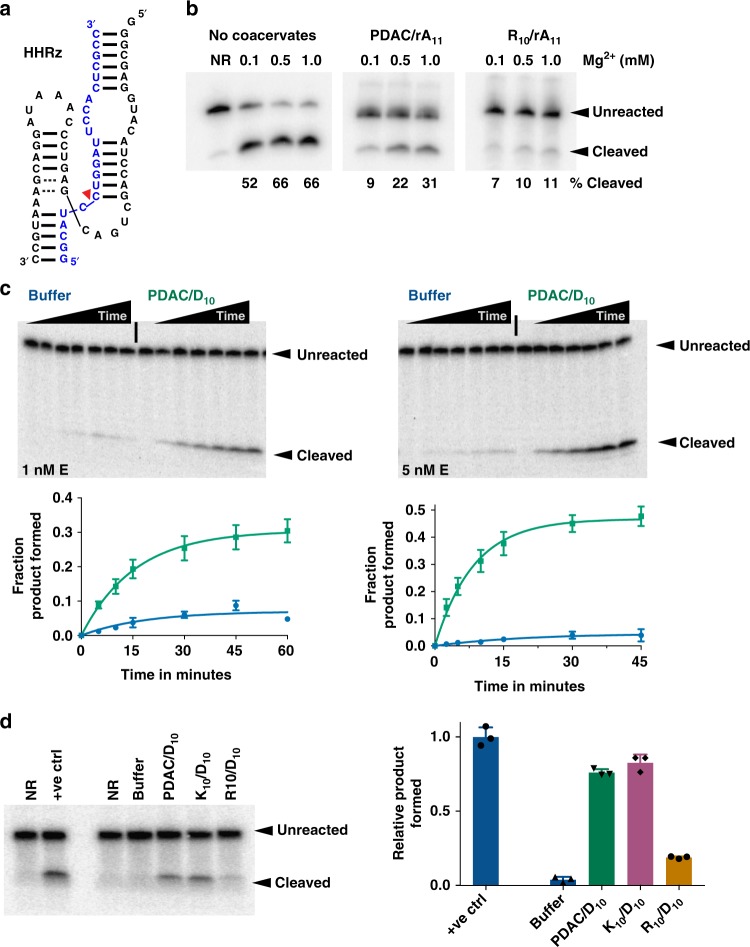
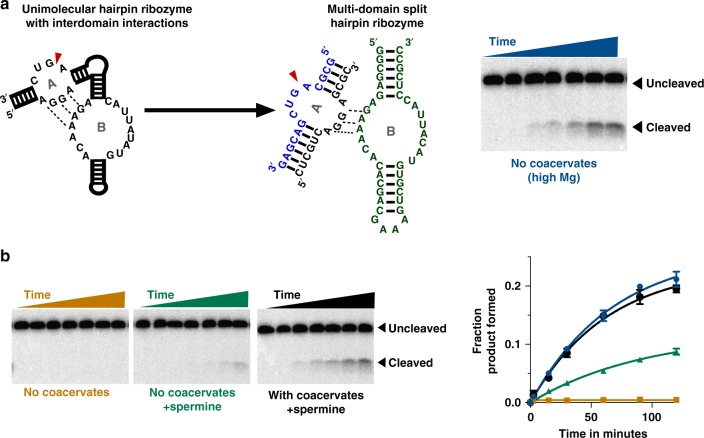
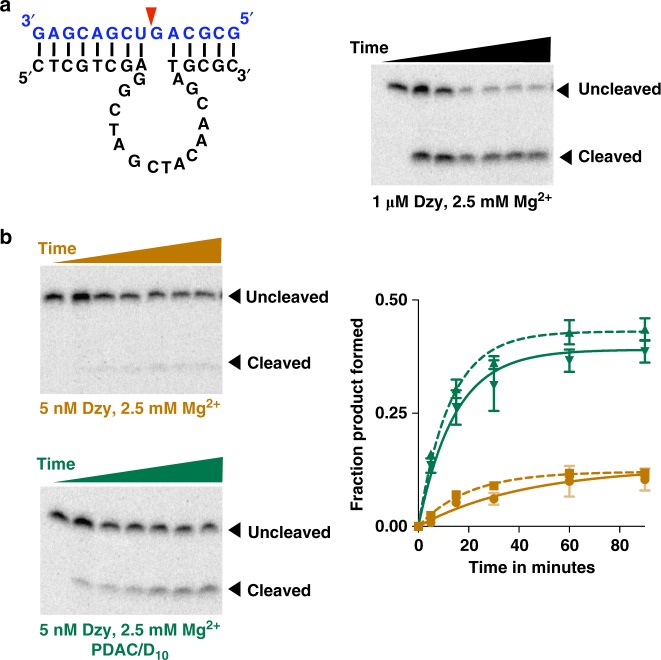
Membraneless compartments, such as complex coacervates, have been hypothesized as plausible prebiotic micro-compartments due to their ability to sequester RNA; however, their compatibility with essential RNA World chemistries is unclear. We show that such compartments can enhance key prebiotically-relevant RNA chemistries. We demonstrate that template-directed RNA polymerization is sensitive to polycation identity, with polydiallyldimethylammonium chloride (PDAC) outperforming poly(allylamine), poly(lysine), and poly(arginine) in polycation/RNA coacervates. Differences in RNA diffusion rates between PDAC/RNA and oligoarginine/RNA coacervates imply distinct biophysical environments. Template-directed RNA polymerization is relatively insensitive to Mg2+ concentration when performed in PDAC/RNA coacervates as compared to buffer, even enabling partial rescue of the reaction in the absence of magnesium. Finally, we show enhanced activities of multiple nucleic acid enzymes including two ribozymes and a deoxyribozyme, underscoring the generality of this approach, in which functional nucleic acids like aptamers and ribozymes, and in some cases key cosolutes localize within the coacervate microenvironments.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
