

Age-related differences in the bone marrow stem cell niche generate specialized microenvironments for the distinct regulation of normal hematopoietic and leukemia stem cells
- PMID: 30700727
- PMCID: PMC6353913
- DOI: 10.1038/s41598-018-36999-5
Age-related differences in the bone marrow stem cell niche generate specialized microenvironments for the distinct regulation of normal hematopoietic and leukemia stem cells
Abstract
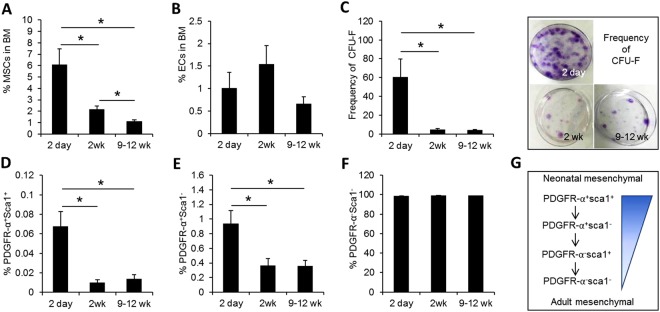
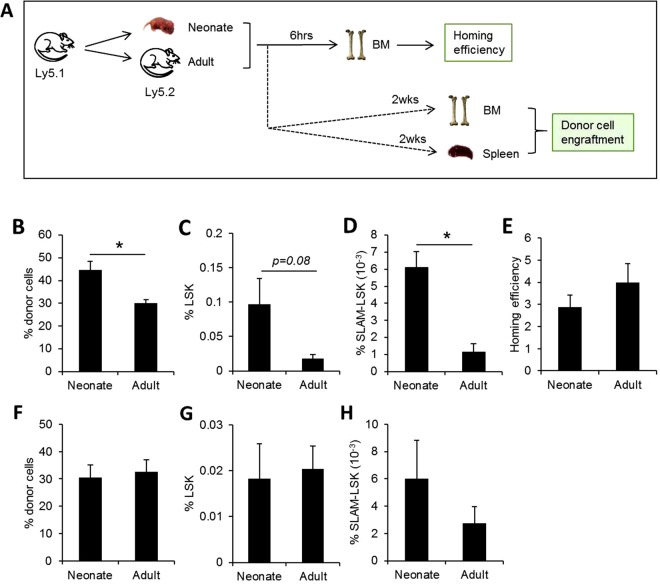
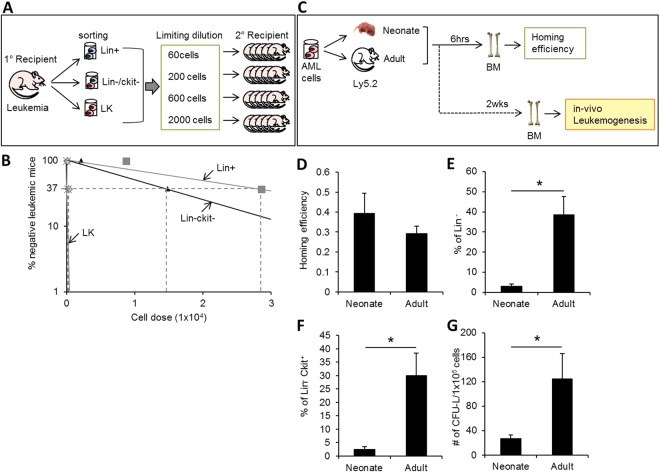
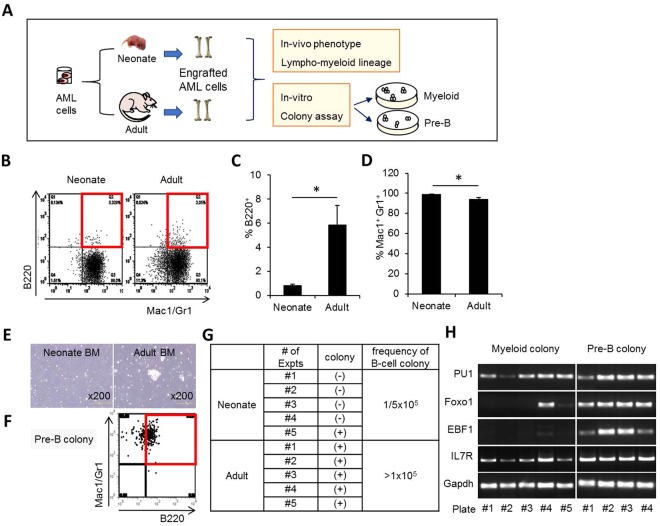
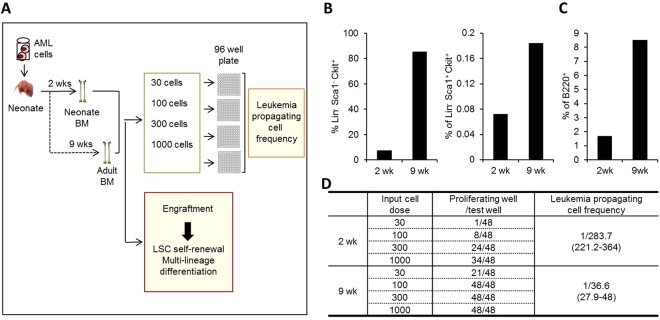
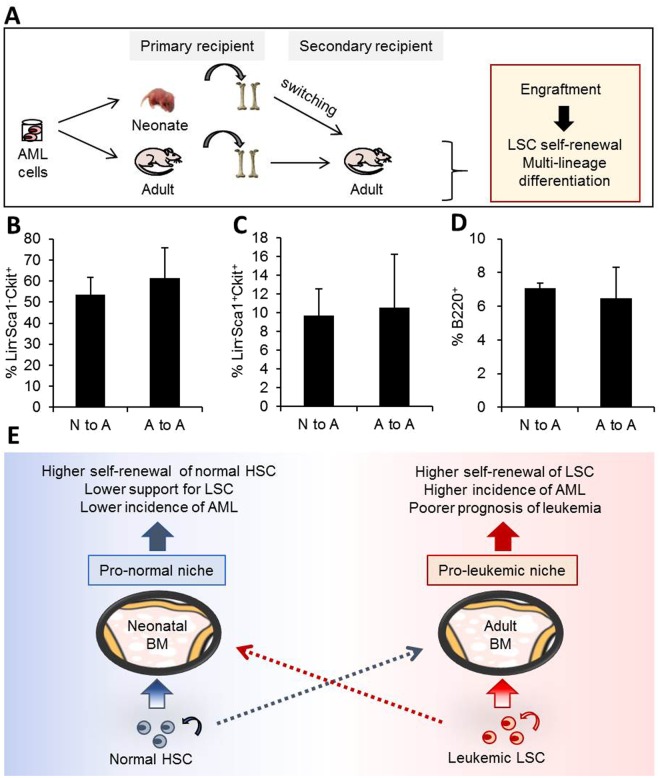
The bone marrow (BM) microenvironment serves as a stem cell niche regulating the in vivo cell fate of normal hematopoietic stem cells (HSC) as well as leukemia stem cells (LSCs). Accumulating studies have indicated that the regeneration of normal HSCs and the process of leukemogenesis change with advancing age. However, the role of microenvironmental factors in these age-related effects are unclear. Here, we compared the stem cell niche in neonatal and adult BM to investigate potential differences in their microenvironmental regulation of both normal and leukemic stem cells. We found that the mesenchymal niche in neonatal BM, compared to adult BM, was characterized by a higher frequency of primitive subsets of mesenchymal stroma expressing both platelet-derived growth factor receptor and Sca-1, and higher expression levels of the niche cross-talk molecules, Jagged-1 and CXCL-12. Accordingly, normal HSCs transplanted into neonatal mice exhibited higher levels of regeneration in BM, with no difference in homing efficiency or splenic engraftment compared to adult BM. In contrast, in vivo self-renewal of LSCs was higher in adult BM than in neonatal BM, with increased frequencies of leukemia-initiating cells as well as higher lympho-myeloid differentiation potential towards biphenotypic leukemic cells. These differences in LSC self-renewal capacity between neonates and adults was abrogated by switching of recipients, confirming their microenvironmental origin. Our study provides insight into the differences in leukemic diseases observed in childhood and adults, and is important for interpretation of many transplantation studies involving neonatal animal models.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
The Adaptive Remodeling of Stem Cell Niche in Stimulated Bone Marrow Counteracts the Leukemic Niche.Stem Cells. 2018 Oct;36(10):1617-1629. doi: 10.1002/stem.2870. Epub 2018 Jul 31. Stem Cells. 2018. PMID: 30004606
-
Microenvironmental remodeling as a parameter and prognostic factor of heterogeneous leukemogenesis in acute myelogenous leukemia.Cancer Res. 2015 Jun 1;75(11):2222-31. doi: 10.1158/0008-5472.CAN-14-3379. Epub 2015 Mar 19. Cancer Res. 2015. PMID: 25791383
-
Leukemia Stem Cells Microenvironment.Adv Exp Med Biol. 2017;1041:19-32. doi: 10.1007/978-3-319-69194-7_3. Adv Exp Med Biol. 2017. PMID: 29204827
-
Regulation of hematopoietic and leukemic stem cells by the immune system.Cell Death Differ. 2015 Feb;22(2):187-98. doi: 10.1038/cdd.2014.89. Epub 2014 Jul 4. Cell Death Differ. 2015. PMID: 24992931 Free PMC article. Review.
-
Where Hematopoietic Stem Cells Live: The Bone Marrow Niche.Antioxid Redox Signal. 2018 Jul 10;29(2):191-204. doi: 10.1089/ars.2017.7419. Epub 2018 Jan 9. Antioxid Redox Signal. 2018. PMID: 29113449 Free PMC article. Review.
Cited by
-
Emerging and Future Targeted Therapies for Pediatric Acute Myeloid Leukemia: Targeting the Leukemia Stem Cells.Biomedicines. 2023 Dec 7;11(12):3248. doi: 10.3390/biomedicines11123248. Biomedicines. 2023. PMID: 38137469 Free PMC article. Review.
-
Insights into the molecular profiles of adult and paediatric acute myeloid leukaemia.Mol Oncol. 2021 Sep;15(9):2253-2272. doi: 10.1002/1878-0261.12899. Epub 2021 Feb 11. Mol Oncol. 2021. PMID: 33421304 Free PMC article. Review.
-
Human Aging Alters the Spatial Organization between CD34+ Hematopoietic Cells and Adipocytes in Bone Marrow.Stem Cell Reports. 2020 Aug 11;15(2):317-325. doi: 10.1016/j.stemcr.2020.06.011. Epub 2020 Jul 9. Stem Cell Reports. 2020. PMID: 32649902 Free PMC article.
-
Common and different alterations of bone marrow mesenchymal stromal cells in myelodysplastic syndrome and multiple myeloma.Cell Prolif. 2020 May;53(5):e12819. doi: 10.1111/cpr.12819. Epub 2020 May 5. Cell Prolif. 2020. PMID: 32372504 Free PMC article.
-
An IL-10/DEL-1 axis supports granulopoiesis and survival from sepsis in early life.Nat Commun. 2024 Jan 23;15(1):680. doi: 10.1038/s41467-023-44178-y. Nat Commun. 2024. PMID: 38263289 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
