

The structural basis of the arrestin binding to GPCRs
- PMID: 30703488
- PMCID: PMC6377262
- DOI: 10.1016/j.mce.2019.01.019
The structural basis of the arrestin binding to GPCRs
Abstract
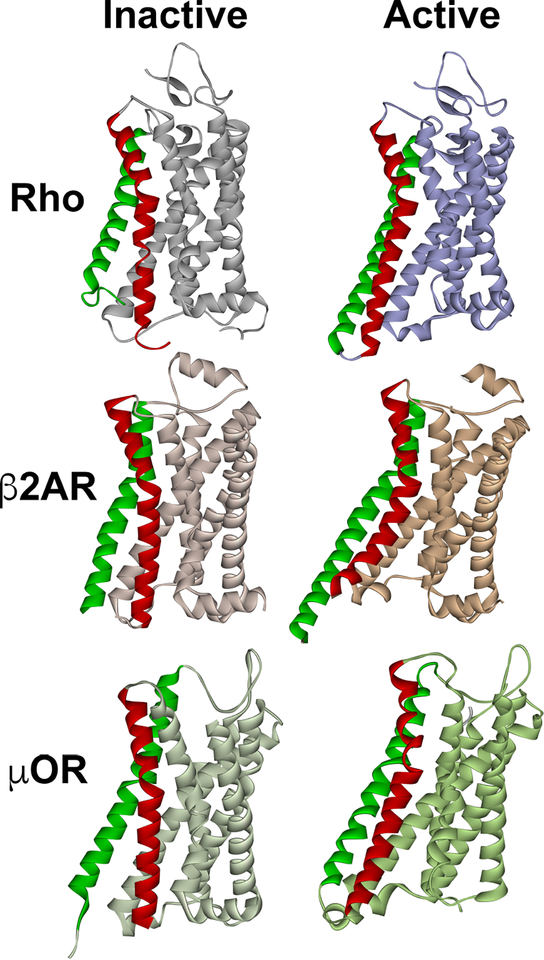
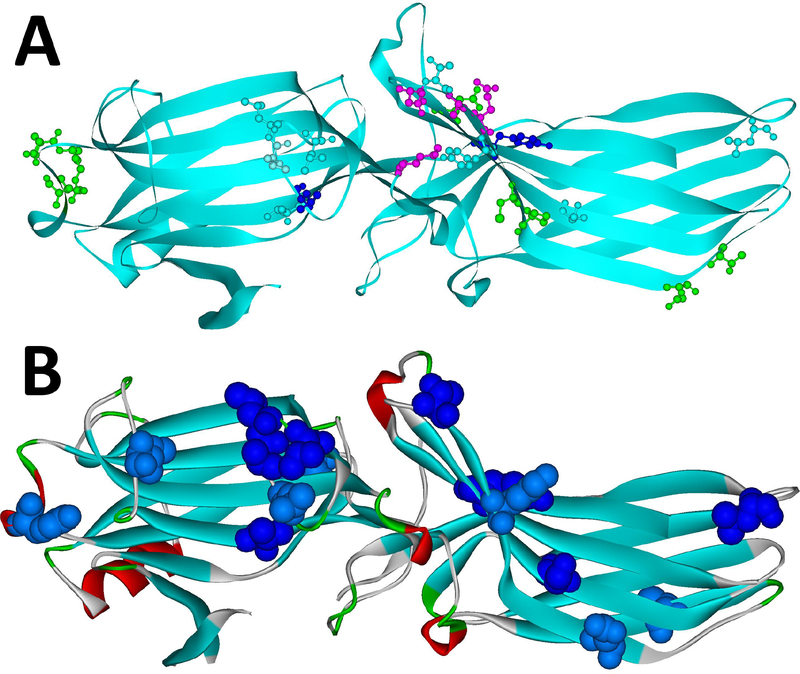
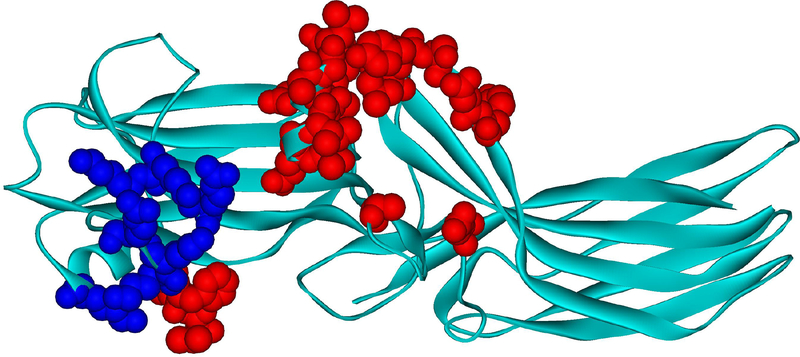
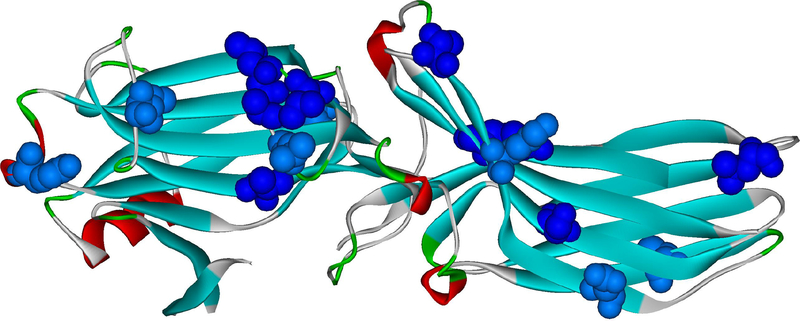
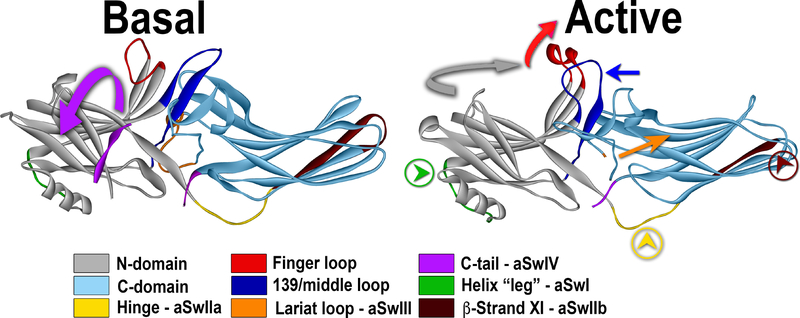
G protein-coupled receptors (GPCRs) are the largest family of signaling proteins targeted by more clinically used drugs than any other protein family. GPCR signaling via G proteins is quenched (desensitized) by the phosphorylation of the active receptor by specific GPCR kinases (GRKs) followed by tight binding of arrestins to active phosphorylated receptors. Thus, arrestins engage two types of receptor elements: those that contain GRK-added phosphates and those that change conformation upon activation. GRKs attach phosphates to serines and threonines in the GPCR C-terminus or any one of the cytoplasmic loops. In addition to these phosphates, arrestins engage the cavity that appears between trans-membrane helices upon receptor activation and several other non-phosphorylated elements. The residues that bind GPCRs are localized on the concave side of both arrestin domains. Arrestins undergo a global conformational change upon receptor binding (become activated). Arrestins serve as important hubs of cellular signaling, emanating from activated GPCRs and receptor-independent.
Keywords: Arrestin; GPCR; Phosphates; Protein engineering; Receptor specificity; Signaling.
Copyright © 2019. Published by Elsevier B.V.
Figures
References
-
- Azevedo AW, Doan T, Moaven H, Sokal I, Baameur F, Vishnivetskiy SA, Homan KT, Tesmer JJ, Gurevich VV, Chen J, et al. (2015). C-terminal threonines and serines play distinct roles in the desensitization of rhodopsin, a G protein-coupled receptor. Elife 4, doi: 10.7554/eLife.05981. - DOI - PMC - PubMed
-
- Baillie GS, Adams DR, Bhari N, Houslay TM, Vadrevu S, Meng D, Li X, Dunlop A, Milligan G, Bolger GB, et al. (2007). Mapping binding sites for the PDE4D5 cAMP-specific phosphodiesterase to the N- and C-domains of beta-arrestin using spot-immobilized peptide arrays. Biochem J 404, 71–80. - PMC - PubMed
-
- Benovic JL, DeBlasi A, Stone WC, Caron MG, and Lefkowitz RJ (1989). Beta-adrenergic receptor kinase: primary structure delineates a multigene family. Science 246, 235–240. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
