

Extranuclear DNA accumulates in aged cells and contributes to senescence and inflammation
- PMID: 30706626
- PMCID: PMC6413746
- DOI: 10.1111/acel.12901
Extranuclear DNA accumulates in aged cells and contributes to senescence and inflammation
Abstract
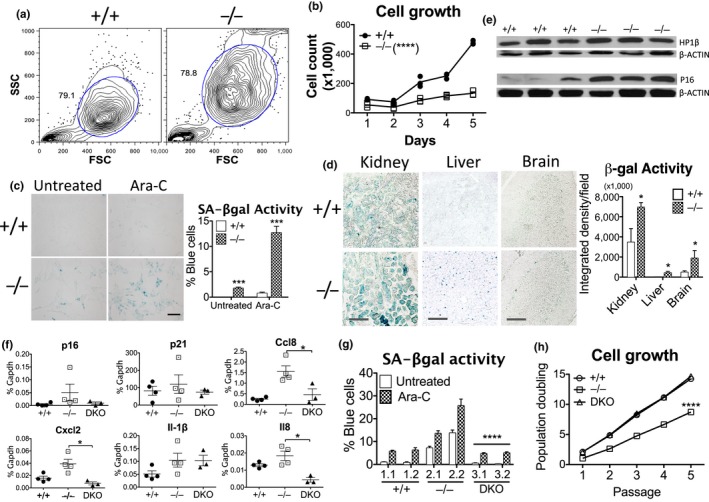
Systemic inflammation is central to aging-related conditions. However, the intrinsic factors that induce inflammation are not well understood. We previously identified a cell-autonomous pathway through which damaged nuclear DNA is trafficked to the cytosol where it activates innate cytosolic DNA sensors that trigger inflammation. These results led us to hypothesize that DNA released after cumulative damage contributes to persistent inflammation in aging cells through a similar mechanism. Consistent with this notion, we found that older cells harbored higher levels of extranuclear DNA compared to younger cells. Extranuclear DNA was exported by a leptomycin B-sensitive process, degraded through the autophagosome-lysosomal pathway and triggered innate immune responses through the DNA-sensing cGAS-STING pathway. Patient cells from the aging diseases ataxia and progeria also displayed extranuclear DNA accumulation, increased pIRF3 and pTBK1, and STING-dependent p16 expression. Removing extranuclear DNA in old cells using DNASE2A reduced innate immune responses and senescence-associated (SA) β-gal enzyme activity. Cells and tissues of Dnase2a-/- mice with defective DNA degradation exhibited slower growth, higher activity of β-gal, or increased expression of HP-1β and p16 proteins, while Dnase2a-/- ;Sting-/- cells and tissues were rescued from these phenotypes, supporting a role for extranuclear DNA in senescence. We hypothesize a direct role for excess DNA in aging-related inflammation and in replicative senescence, and propose DNA degradation as a therapeutic approach to remove intrinsic DNA and revert inflammation associated with aging.
Keywords: Dnase2a; STING pathway; cellular senescence; extranuclear DNA; inflammation; premature aging.
© 2019 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Coppé, J. P. , Patil, C. K. , Rodier, F. , Krtolica, A. , Beausejour, C. M. , Parrinello, S. , … Campisi, J. (2010). A human‐like senescence‐associated secretory phenotype is conserved in mouse cells dependent on physiological oxygen. PLoS One, 5(2), e9188 10.1371/journal.pone.0009188. - DOI - PMC - PubMed
-
- De Vos, W. H. , Houben, F. , Kamps, M. , Malhas, A. , Verheyen, F. , Cox, J. , … Broers, J. L. (2011). Repetitive disruptions of the nuclear envelope invoke temporary loss of cellular compartmentalization in laminopathies. Human Molecular Genetics, 20(21), 4175–4186. 10.1093/hmg/ddr344. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
