

Identification of the X-linked germ cell specific miRNAs (XmiRs) and their functions
- PMID: 30707741
- PMCID: PMC6358104
- DOI: 10.1371/journal.pone.0211739
Identification of the X-linked germ cell specific miRNAs (XmiRs) and their functions
Abstract
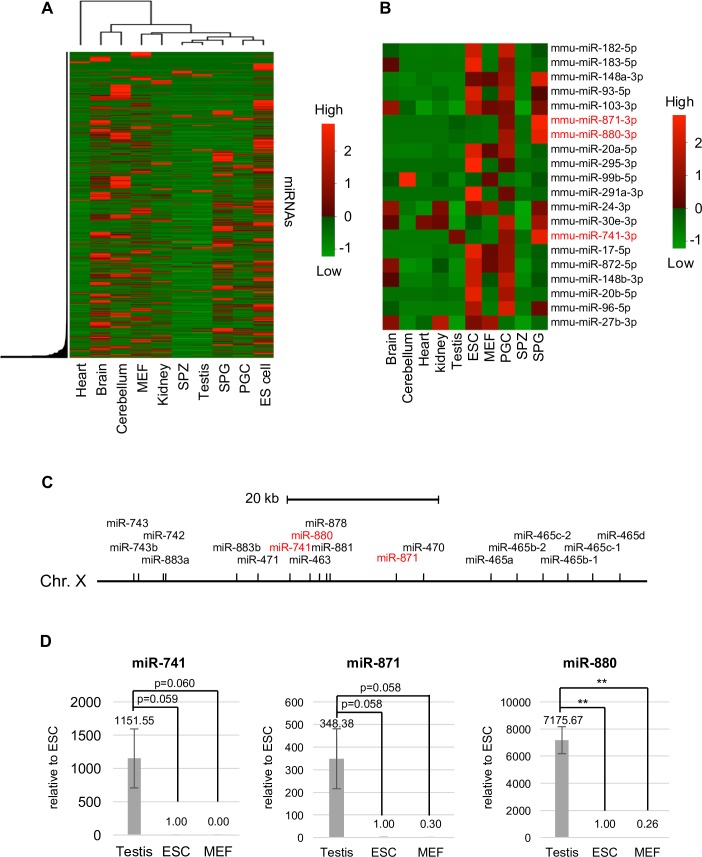
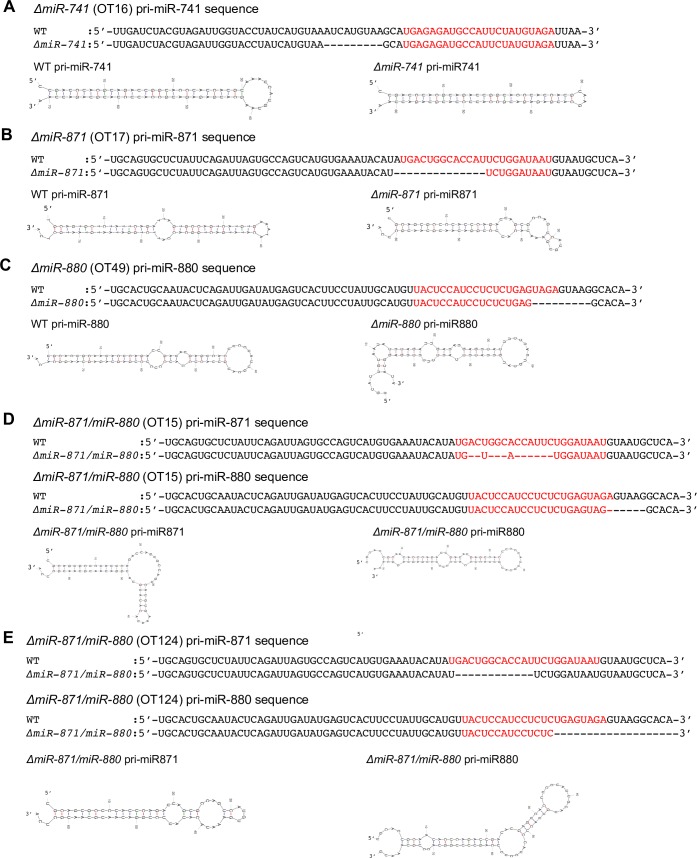
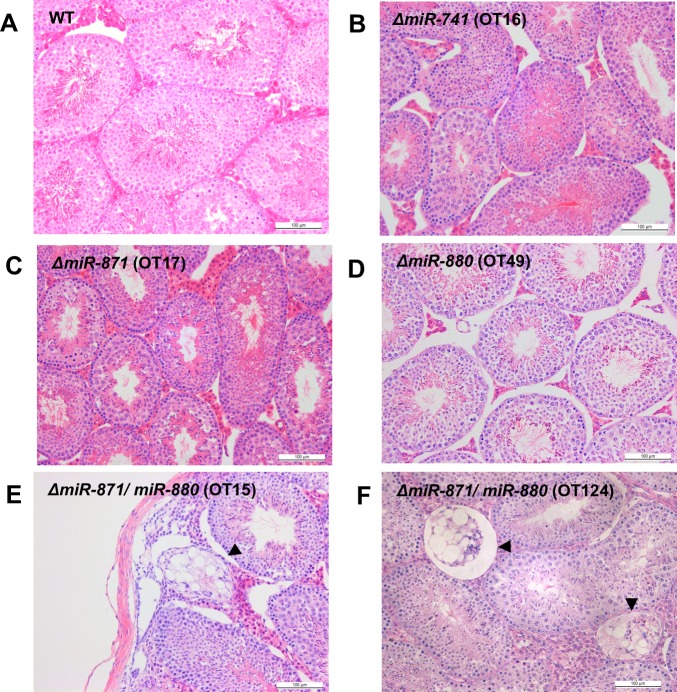
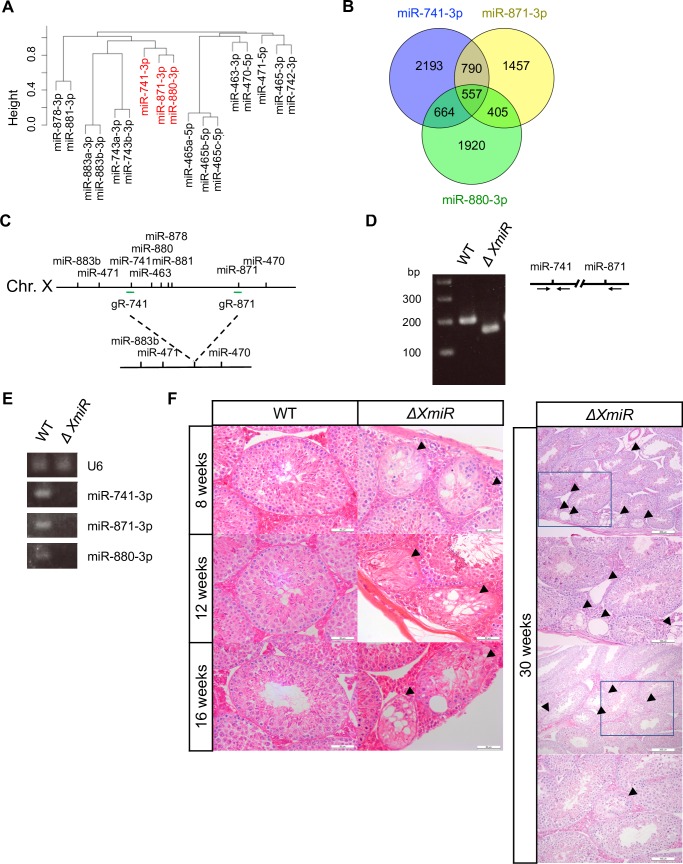
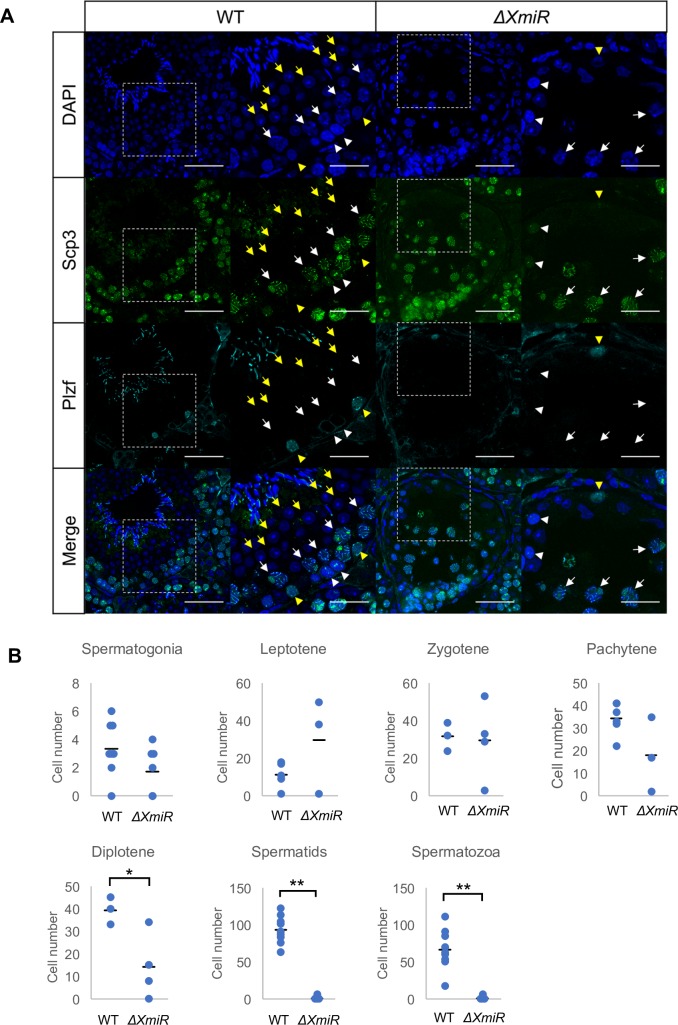
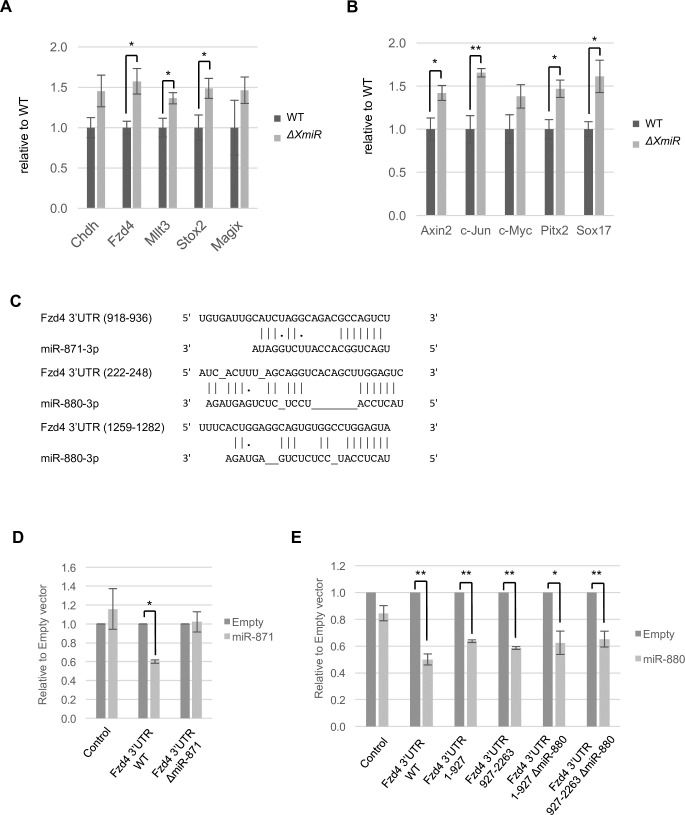
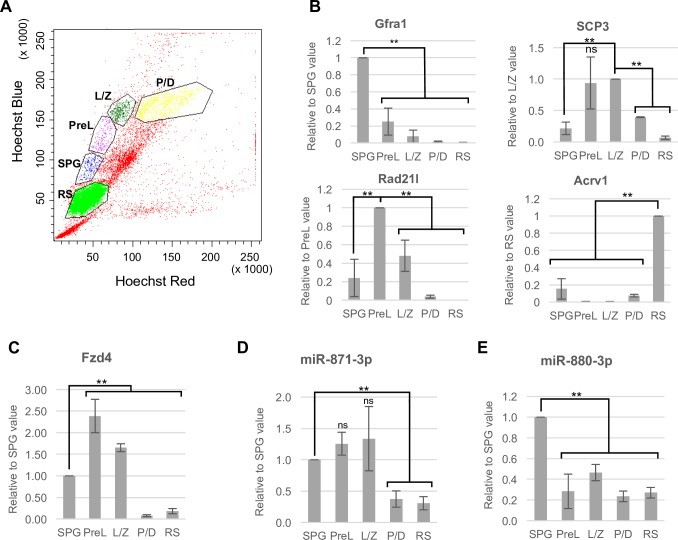
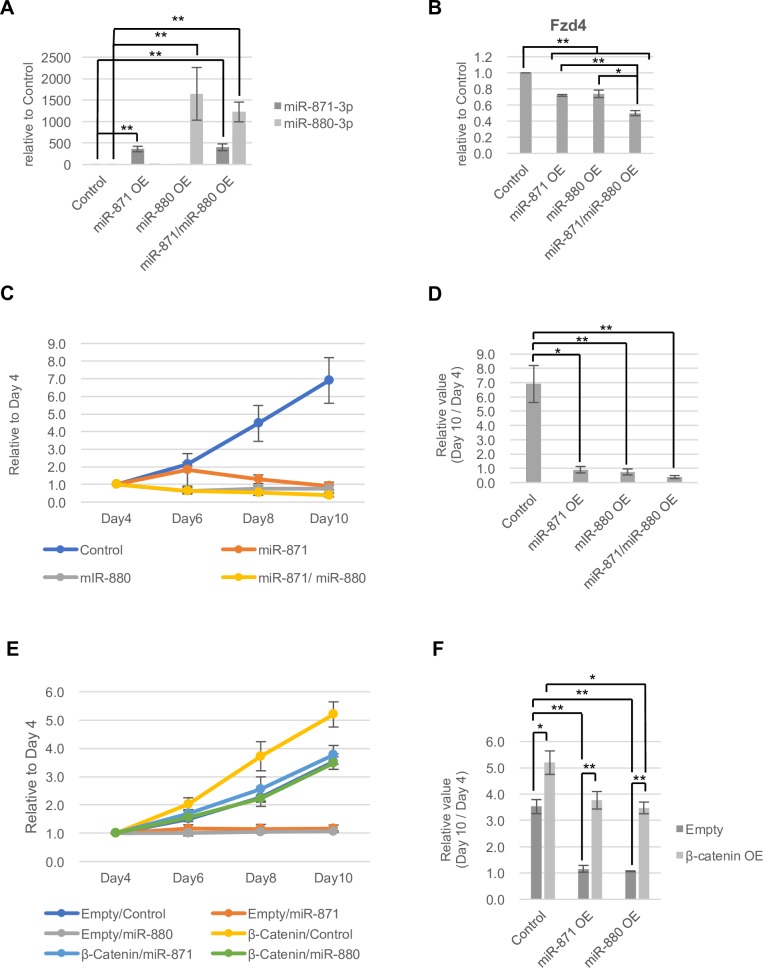
MicroRNAs (miRNAs) play a critical role in multiple aspects of biology. Dicer, an RNase III endonuclease, is essential for the biogenesis of miRNAs, and the germ cell-specific Dicer1 knockout mouse shows severe defects in gametogenesis. How miRNAs regulate germ cell development is still not fully understood. In this study, we identified germ cell-specific miRNAs (miR-741-3p, miR-871-3p, miR-880-3p) by analyzing published RNA-seq data of mouse. These miRNA genes are contiguously located on the X chromosome near other miRNA genes. We named them X chromosome-linked miRNAs (XmiRs). To elucidate the functions of XmiRs, we generated knockout mice of these miRNA genes using the CRISPR/Cas9-mediated genome editing system. Although no histological abnormalities were observed in testes of F0 mice in which each miRNA gene was disrupted, a deletion covering miR-871 and miR-880 or covering all XmiRs (ΔXmiRs) resulted in arrested spermatogenesis in meiosis in a few seminiferous tubules, indicating their redundant functions in spermatogenesis. Among candidate targets of XmiRs, we found increased expression of a gene encoding a WNT receptor, FZD4, in ΔXmiRs testis compared with that in wildtype testis. miR-871-3p and miR-880-3p repressed the expression of Fzd4 via the 3'-untranslated region of its mRNA. In addition, downstream genes of the WNT/β-catenin pathway were upregulated in ΔXmiRs testis. We also found that miR-871, miR-880, and Fzd4 were expressed in spermatogonia, spermatocytes and spermatids, and overexpression of miR-871 and miR-880 in germ stem cells in culture repressed their increase in number and Fzd4 expression. Previous studies indicated that the WNT/β-catenin pathway enhances and represses proliferation and differentiation of spermatogonia, respectively, and our results consistently showed that stable β-catenin enhanced GSC number. In addition, stable β-catenin partially rescued reduced GSC number by overexpression of miR-871 and miR-880. The results together suggest that miR-871 and miR-880 cooperatively regulate the WNT/β-catenin pathway during testicular germ cell development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Ginsburg M, Snow MH, McLaren A. Primordial germ cells in the mouse embryo during gastrulation. Development. 1990;110(2):521–8. - PubMed
-
- McLaren A. Meiosis and differentiation of mouse germ cells. Symp Soc Exp Biol. 1984;38:7–23. - PubMed
-
- de Rooij DG. Proliferation and differentiation of spermatogonial stem cells. Reproduction. 2001;121(3):347–54. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
