

Control of the Antitumor Immune Response by Cancer Metabolism
- PMID: 30708988
- PMCID: PMC6406288
- DOI: 10.3390/cells8020104
Control of the Antitumor Immune Response by Cancer Metabolism
Abstract
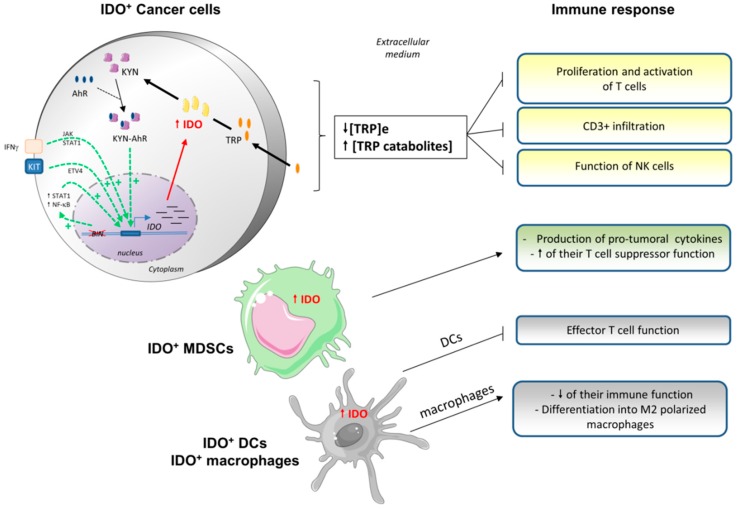
The metabolic reprogramming of tumor cells and immune escape are two major hallmarks of cancer cells. The metabolic changes that occur during tumorigenesis, enabling survival and proliferation, are described for both solid and hematological malignancies. Concurrently, tumor cells have deployed mechanisms to escape immune cell recognition and destruction. Additionally, therapeutic blocking of tumor-mediated immunosuppression has proven to have an unprecedented positive impact in clinical oncology. Increased evidence suggests that cancer metabolism not only plays a crucial role in cancer signaling for sustaining tumorigenesis and survival, but also has wider implications in the regulation of antitumor immune signaling through both the release of signaling molecules and the expression of immune membrane ligands. Here, we review these molecular events to highlight the contribution of cancer cell metabolic reprogramming on the shaping of the antitumor immune response.
Keywords: cancer; immunity; metabolism.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
