

Activity Correlations between Direction-Selective Retinal Ganglion Cells Synergistically Enhance Motion Decoding from Complex Visual Scenes
- PMID: 30709656
- PMCID: PMC6424814
- DOI: 10.1016/j.neuron.2019.01.003
Activity Correlations between Direction-Selective Retinal Ganglion Cells Synergistically Enhance Motion Decoding from Complex Visual Scenes
Abstract
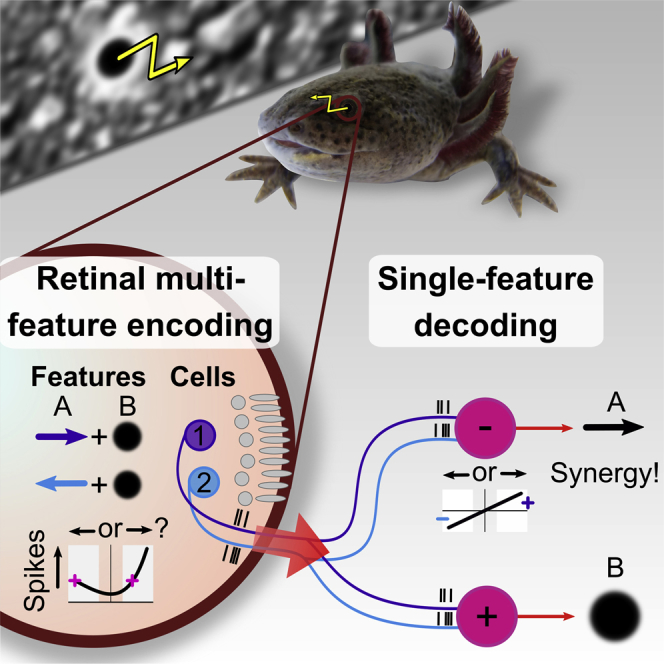
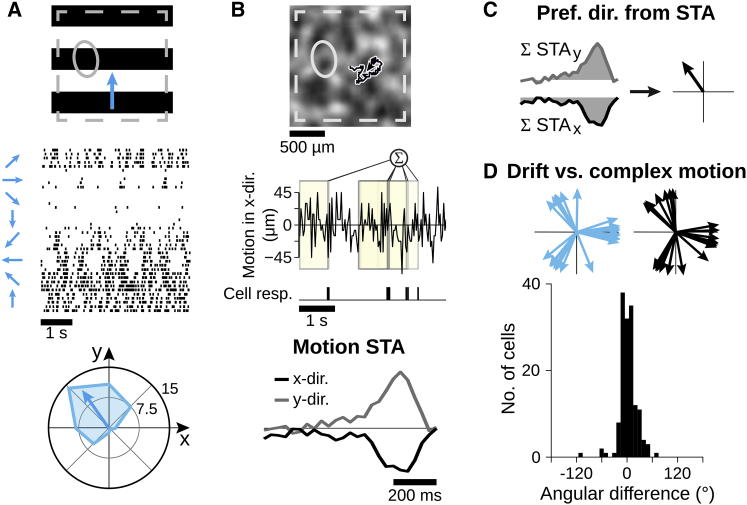
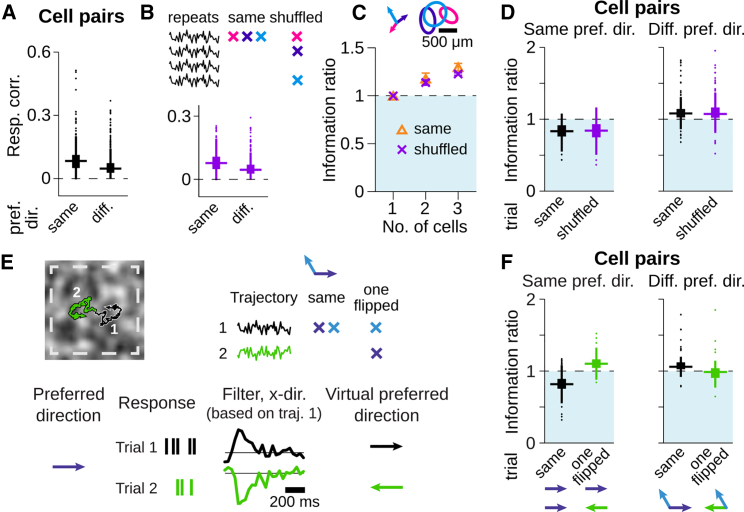
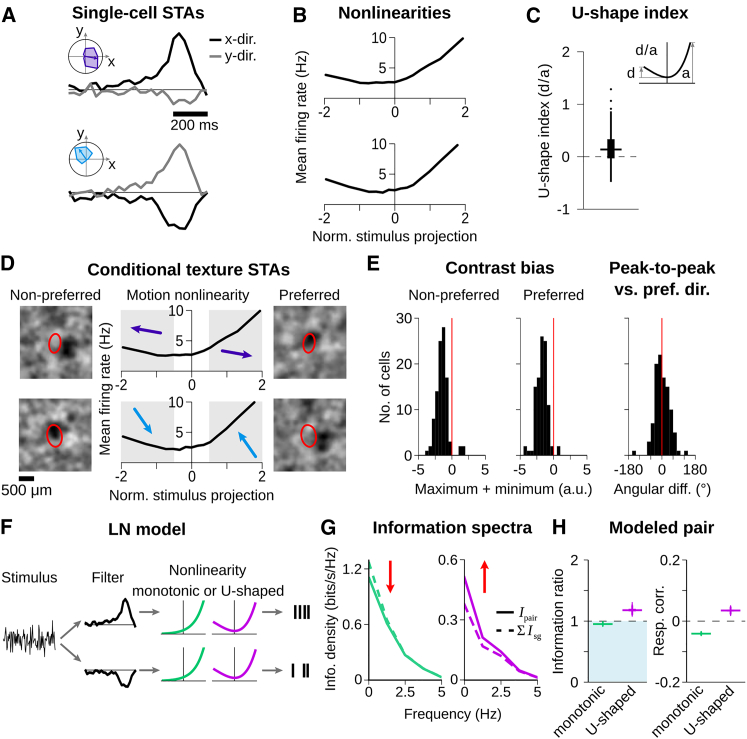
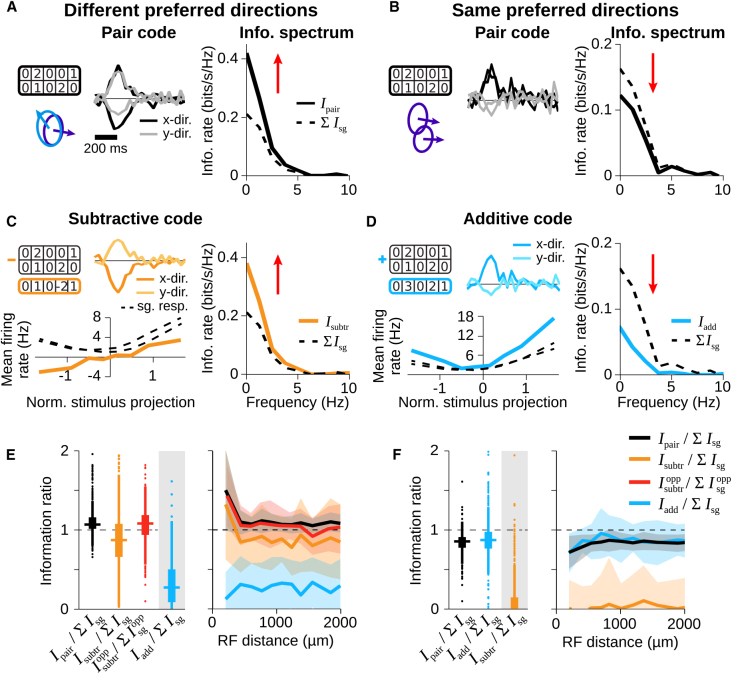
Neurons in sensory systems are often tuned to particular stimulus features. During complex naturalistic stimulation, however, multiple features may simultaneously affect neuronal responses, which complicates the readout of individual features. To investigate feature representation under complex stimulation, we studied how direction-selective ganglion cells in salamander retina respond to texture motion where direction, velocity, and spatial pattern inside the receptive field continuously change. We found that the cells preserve their direction preference under this stimulation, yet their direction encoding becomes ambiguous due to simultaneous activation by luminance changes. The ambiguities can be resolved by considering populations of direction-selective cells with different preferred directions. This gives rise to synergistic motion decoding, yielding more information from the population than the summed information from single-cell responses. Strong positive response correlations between cells with different preferred directions amplify this synergy. Our results show how correlated population activity can enhance feature extraction in complex visual scenes.
Keywords: correlations; direction selectivity; feature extraction; linear decoder; model; population code; retinal ganglion cells; synergy; texture motion; vision.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Figures




References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
