

Chromatin architecture reorganization during neuronal cell differentiation in Drosophila genome
- PMID: 30709849
- PMCID: PMC6442379
- DOI: 10.1101/gr.246710.118
Chromatin architecture reorganization during neuronal cell differentiation in Drosophila genome
Abstract
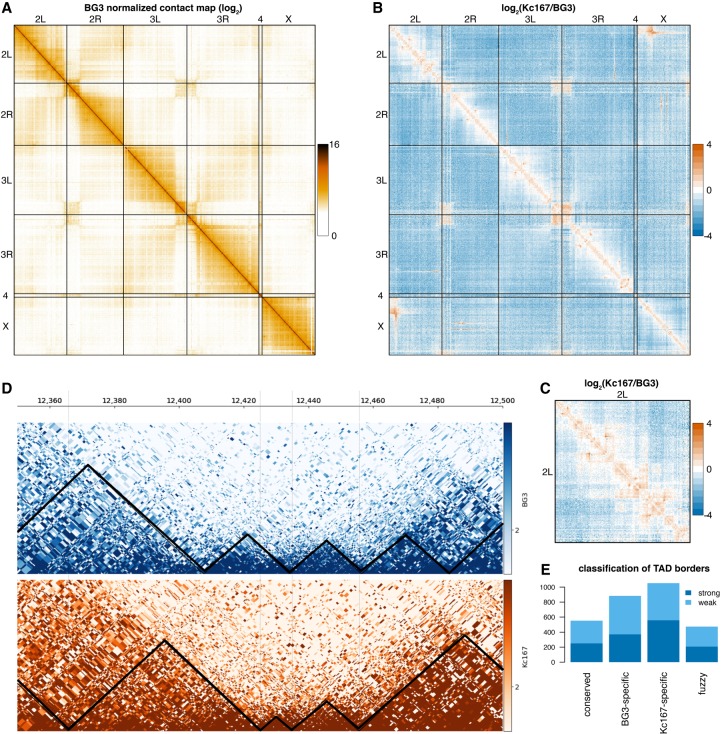
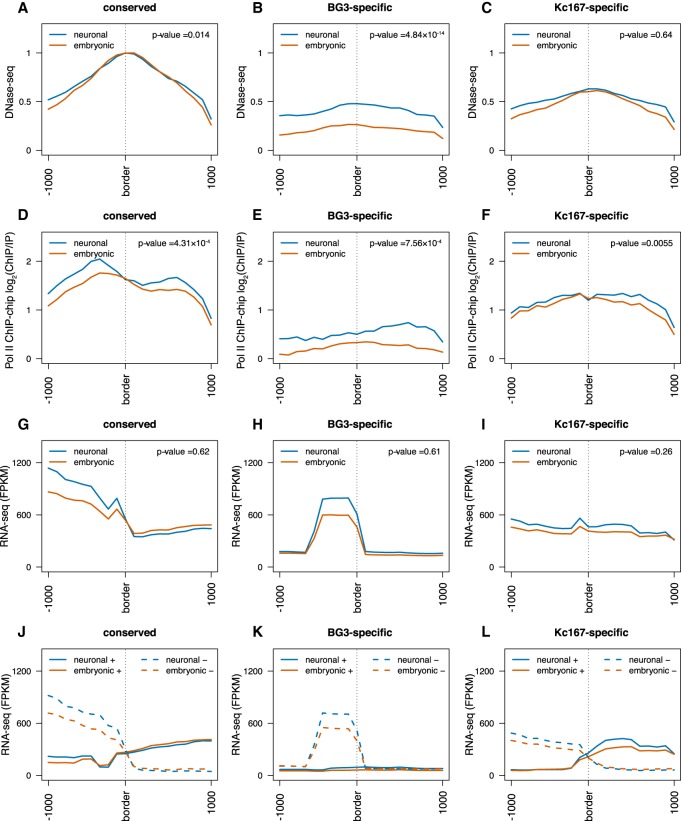
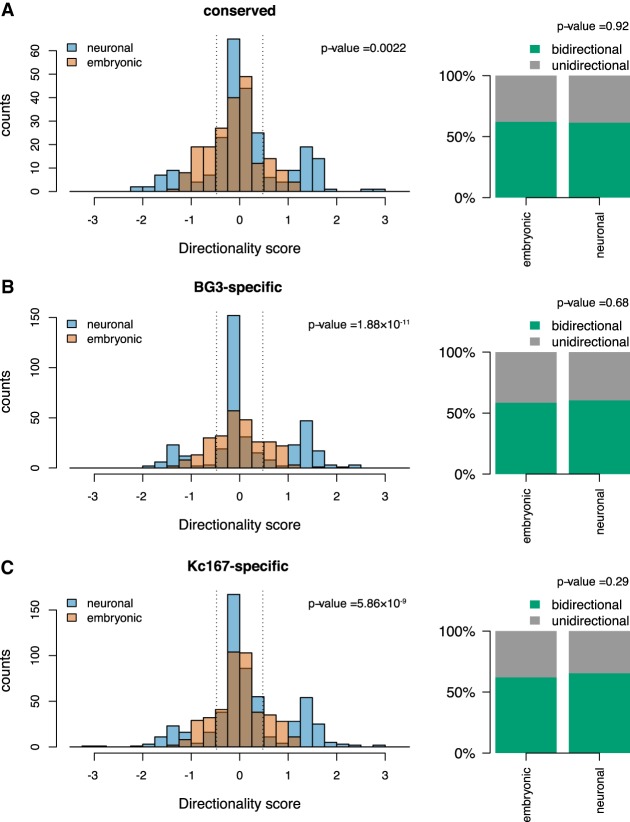
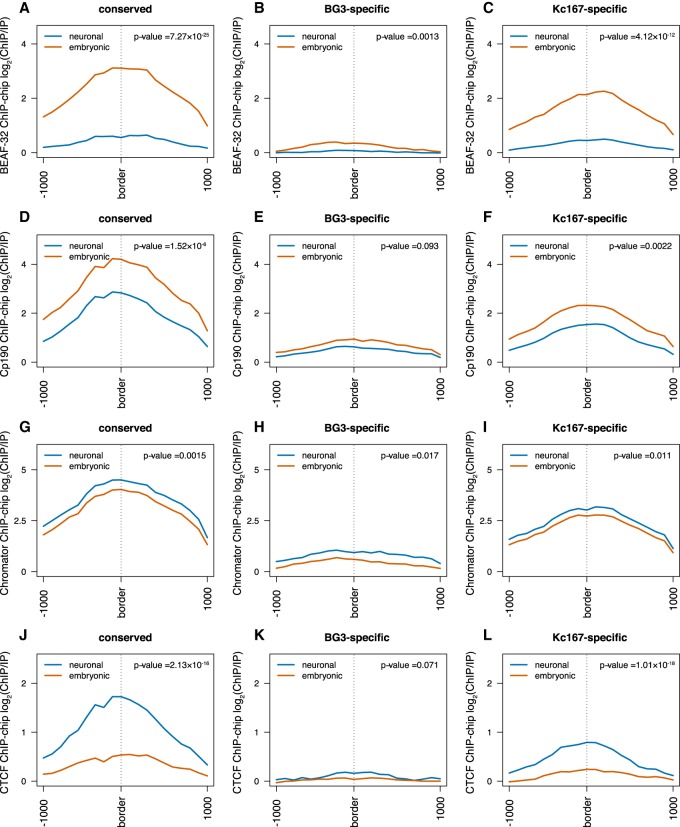
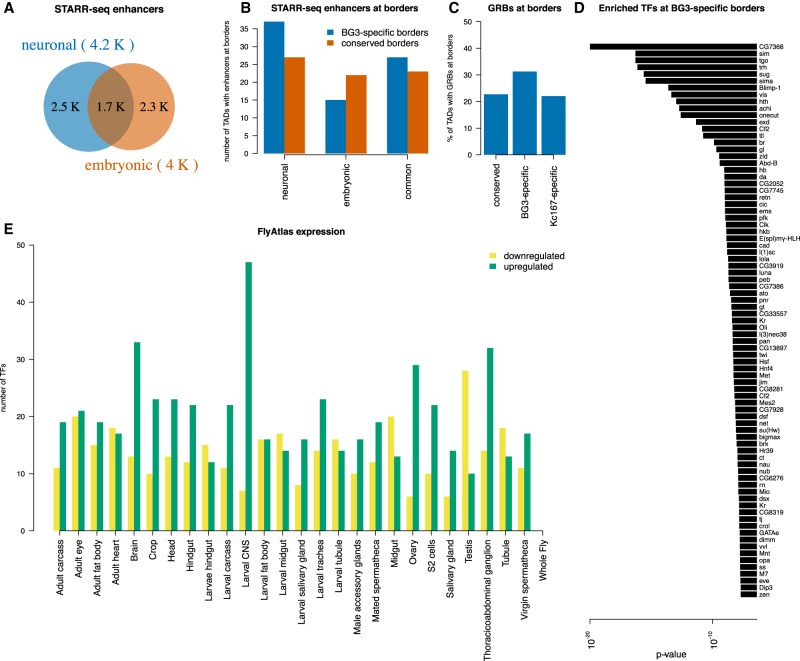
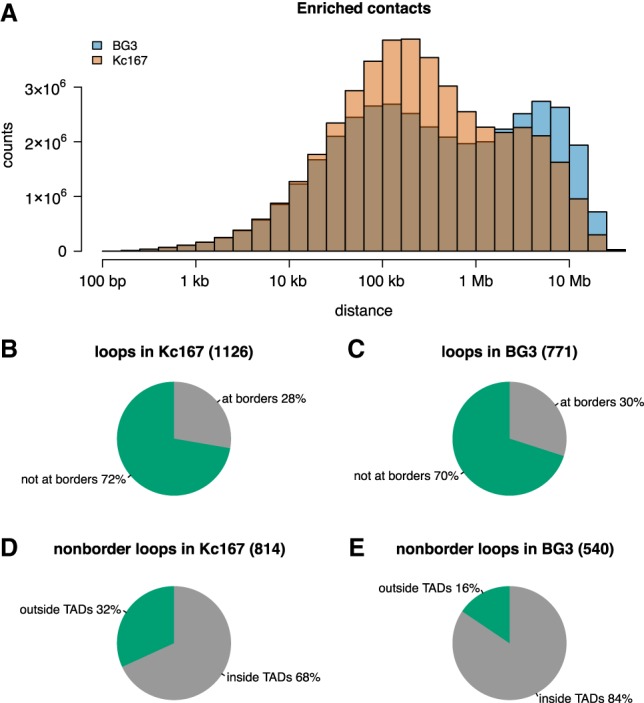
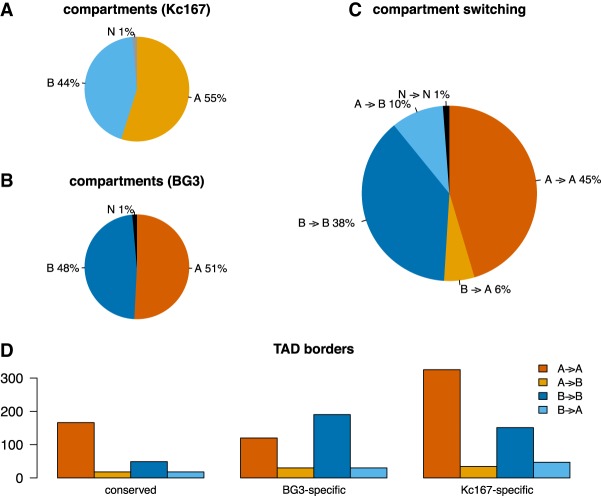
The organization of the genome into topologically associating domains (TADs) was shown to have a regulatory role in development and cellular function, but the mechanism involved in TAD establishment is still unclear. Here, we present the first high-resolution contact map of Drosophila neuronal cells (BG3) and identify different classes of TADs by comparing this to genome organization in embryonic cells (Kc167). We find that only some TADs are conserved in both cell lines, whereas the rest are cell-type-specific. This is supported by a change in the enrichment of architectural proteins at TAD borders, with BEAF-32 present in embryonic cells and CTCF in neuronal cells. Furthermore, we observe strong divergent transcription, together with RNA Polymerase II occupancy and an increase in DNA accessibility at the TAD borders. TAD borders that are specific to neuronal cells are enriched in enhancers controlled by neuronal-specific transcription factors. Our results suggest that TADs are dynamic across developmental stages and reflect the interplay between insulators, transcriptional states, and enhancer activities.
© 2019 Chathoth and Zabet; Published by Cold Spring Harbor Laboratory Press.
Figures
Similar articles
-
Sub-kb Hi-C in D. melanogaster reveals conserved characteristics of TADs between insect and mammalian cells.Nat Commun. 2018 Jan 15;9(1):188. doi: 10.1038/s41467-017-02526-9. Nat Commun. 2018. PMID: 29335463 Free PMC article.
-
High-resolution TADs reveal DNA sequences underlying genome organization in flies.Nat Commun. 2018 Jan 15;9(1):189. doi: 10.1038/s41467-017-02525-w. Nat Commun. 2018. PMID: 29335486 Free PMC article.
-
Active chromatin and transcription play a key role in chromosome partitioning into topologically associating domains.Genome Res. 2016 Jan;26(1):70-84. doi: 10.1101/gr.196006.115. Epub 2015 Oct 30. Genome Res. 2016. PMID: 26518482 Free PMC article.
-
Chromatin Architecture in the Fly: Living without CTCF/Cohesin Loop Extrusion?: Alternating Chromatin States Provide a Basis for Domain Architecture in Drosophila.Bioessays. 2019 Sep;41(9):e1900048. doi: 10.1002/bies.201900048. Epub 2019 Jul 1. Bioessays. 2019. PMID: 31264253 Review.
-
TADs and Their Borders: Free Movement or Building a Wall?J Mol Biol. 2020 Feb 7;432(3):643-652. doi: 10.1016/j.jmb.2019.11.025. Epub 2019 Dec 27. J Mol Biol. 2020. PMID: 31887284 Review.
Cited by
-
CTCF As an Example of DNA-Binding Transcription Factors Containing Clusters of C2H2-Type Zinc Fingers.Acta Naturae. 2021 Jan-Mar;13(1):31-46. doi: 10.32607/actanaturae.11206. Acta Naturae. 2021. PMID: 33959385 Free PMC article.
-
A global high-density chromatin interaction network reveals functional long-range and trans-chromosomal relationships.Genome Biol. 2022 Nov 9;23(1):238. doi: 10.1186/s13059-022-02790-z. Genome Biol. 2022. PMID: 36352464 Free PMC article.
-
Mechanisms of Enhancer-Promoter Interactions in Higher Eukaryotes.Int J Mol Sci. 2021 Jan 12;22(2):671. doi: 10.3390/ijms22020671. Int J Mol Sci. 2021. PMID: 33445415 Free PMC article. Review.
-
Insulator BEAF32 regulates expression of tissue-specific genes and piRNA source loci in Drosophila ovaries.Epigenetics Chromatin. 2025 Jul 28;18(1):49. doi: 10.1186/s13072-025-00613-6. Epigenetics Chromatin. 2025. PMID: 40717082 Free PMC article.
-
Independence of chromatin conformation and gene regulation during Drosophila dorsoventral patterning.Nat Genet. 2021 Apr;53(4):487-499. doi: 10.1038/s41588-021-00799-x. Epub 2021 Apr 1. Nat Genet. 2021. PMID: 33795866 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases