

A Murine Model of Chronic Lymphocytic Leukemia Based on B Cell-Restricted Expression of Sf3b1 Mutation and Atm Deletion
- PMID: 30712845
- PMCID: PMC6372356
- DOI: 10.1016/j.ccell.2018.12.013
A Murine Model of Chronic Lymphocytic Leukemia Based on B Cell-Restricted Expression of Sf3b1 Mutation and Atm Deletion
Abstract
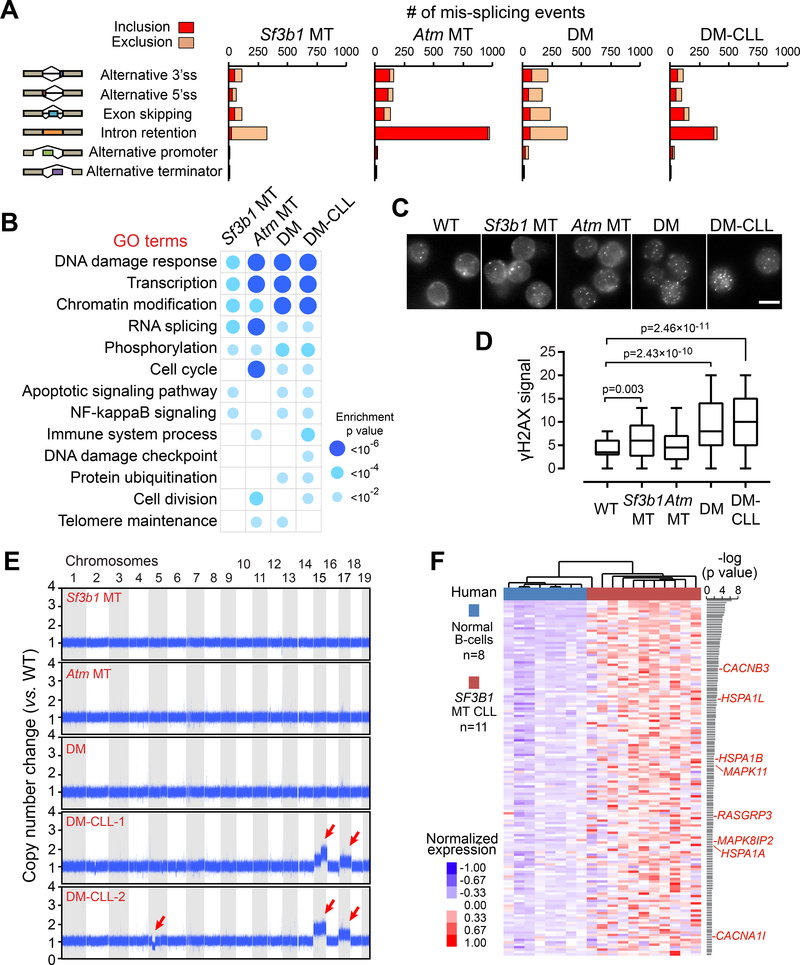
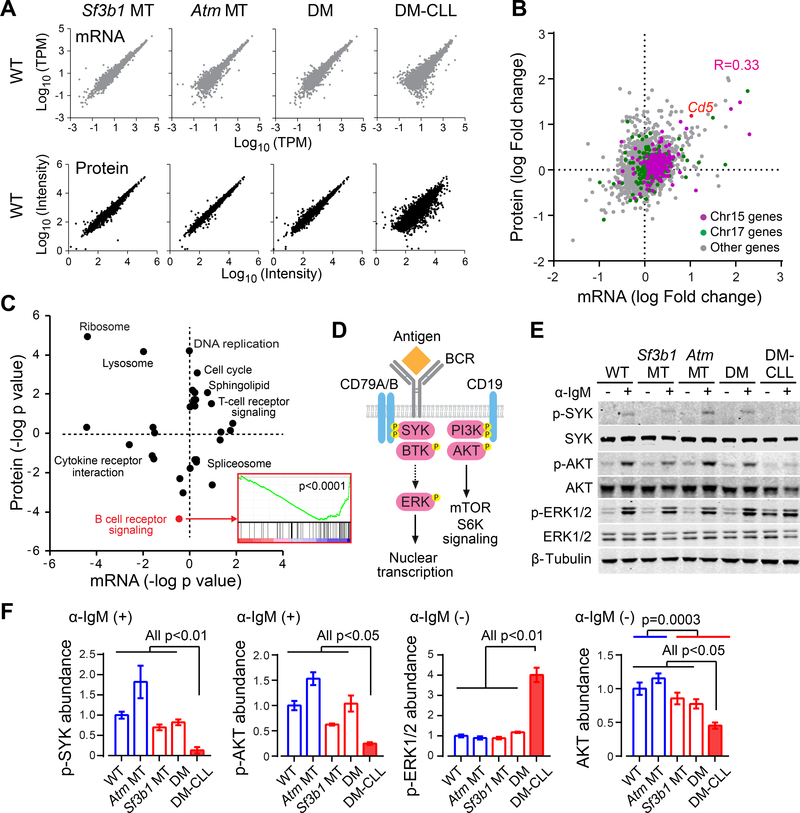
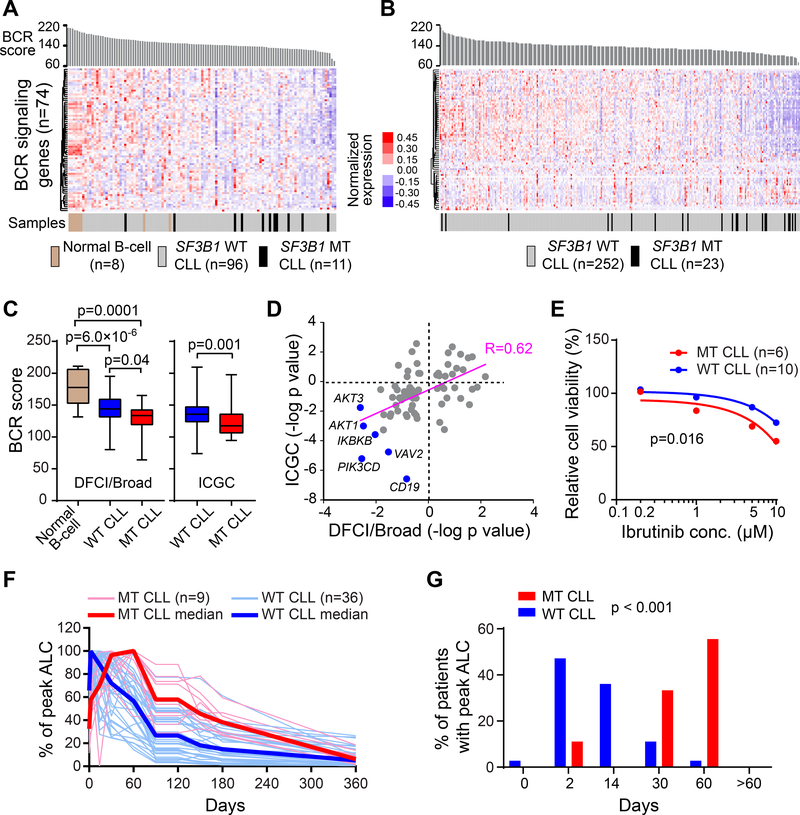
SF3B1 is recurrently mutated in chronic lymphocytic leukemia (CLL), but its role in the pathogenesis of CLL remains elusive. Here, we show that conditional expression of Sf3b1-K700E mutation in mouse B cells disrupts pre-mRNA splicing, alters cell development, and induces a state of cellular senescence. Combination with Atm deletion leads to the overcoming of cellular senescence and the development of CLL-like disease in elderly mice. These CLL-like cells show genome instability and dysregulation of multiple CLL-associated cellular processes, including deregulated B cell receptor signaling, which we also identified in human CLL cases. Notably, human CLLs harboring SF3B1 mutations exhibit altered response to BTK inhibition. Our murine model of CLL thus provides insights into human CLL disease mechanisms and treatment.
Keywords: ATM; BCR signaling; CLL; SF3B1; murine model.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
C.J.W. is a co-founder of Neon therapeutics. C.J.W., D.N. and G.G. receive research funding from Pharmacyclics. B.L.E. has been a consultant for H3 Biomedicine and received research funding from Celgene. G.G receives research funds from IBM. G.G. is an inventor on patent applications related to MuTect, ABSOLUTE and other bioinformatics methods. J.A.D. has received honoraria for participation in advisory board from Merck. J.A.D. has received research funding from Constellation Pharmaceuticals. J.S. is a current employee of Moderna Therapeutics. All other authors do not have any relevant conflict of interest.
Figures
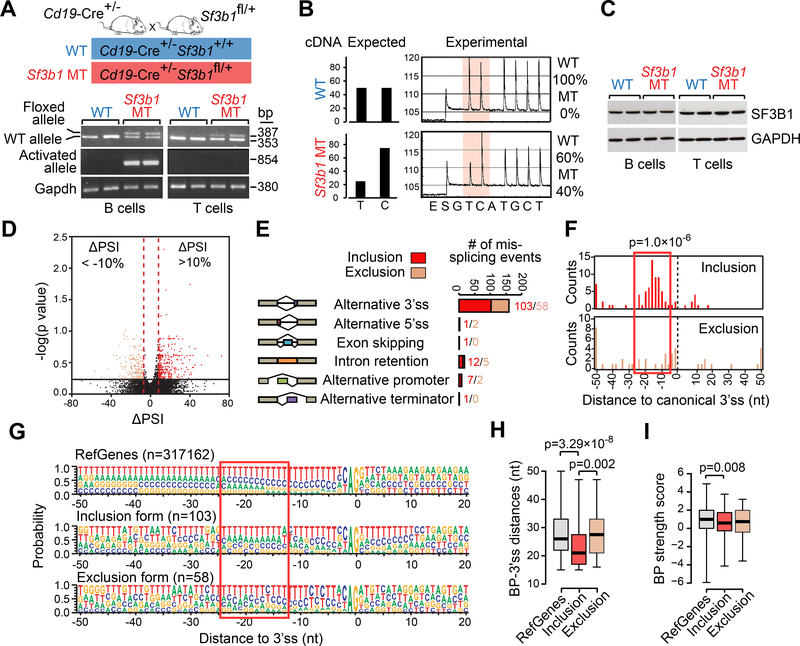
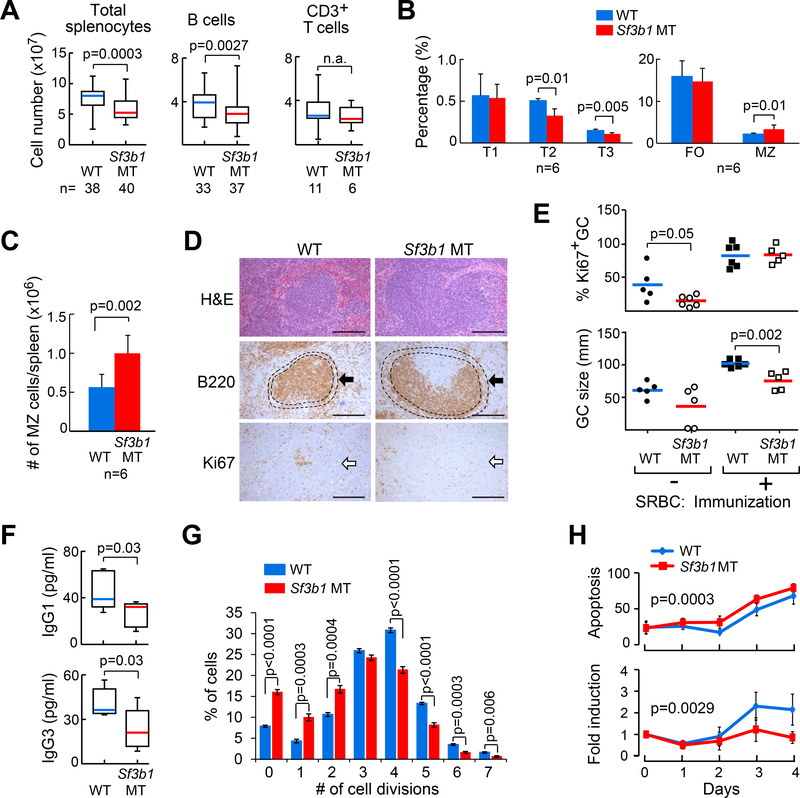
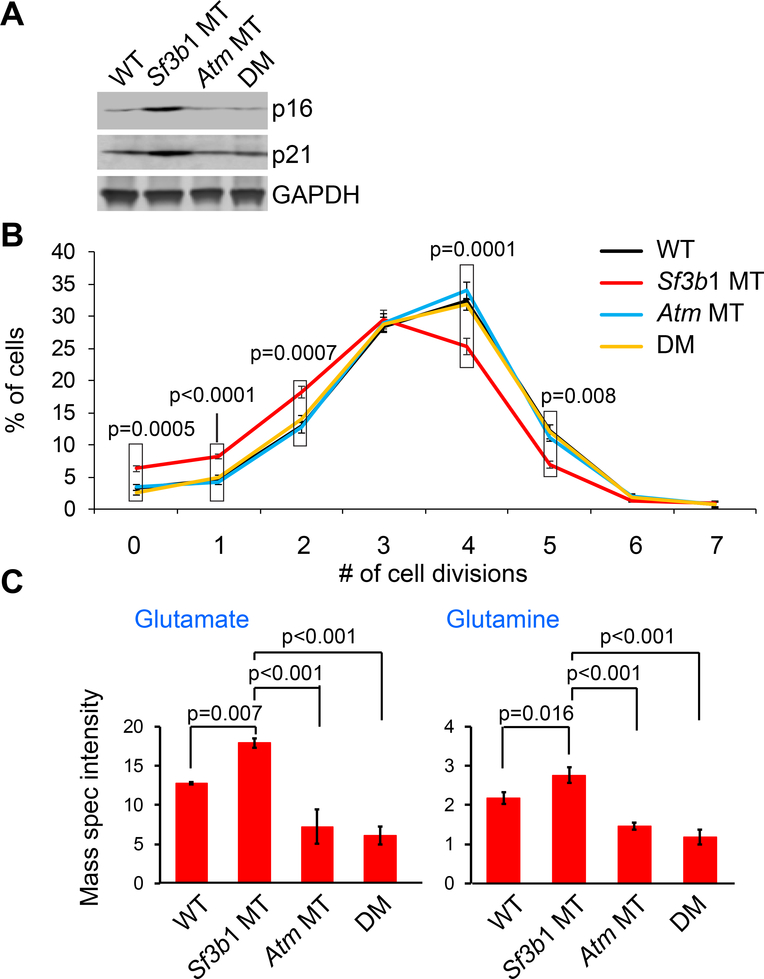
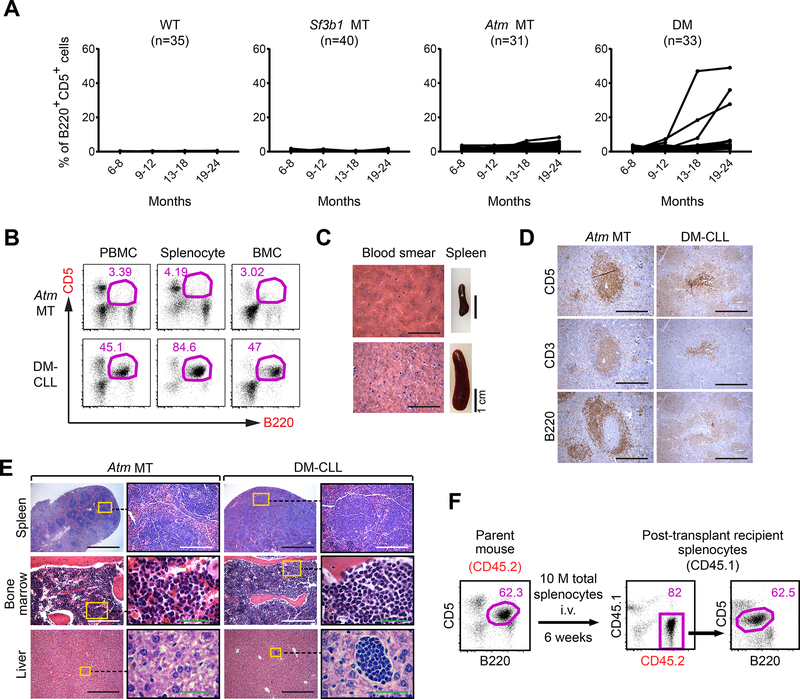
References
-
- Alfarano A, Indraccolo S, Circosta P, Minuzzo S, Vallario A, Zamarchi R, Fregonese A, Calderazzo F, Faldella A, Aragno M, et al. (1999). An alternatively spliced form of CD79b gene may account for altered B-cell receptor expression in B-chronic lymphocytic leukemia. Blood 93, 2327–2335. - PubMed
-
- Bartkova J, Rezaei N, Liontos M, Karakaidos P, Kletsas D, Issaeva N, Vassiliou LV, Kolettas E, Niforou K, Zoumpourlis VC, et al. (2006). Oncogene-induced senescence is part of the tumorigenesis barrier imposed by DNA damage checkpoints. Nature 444, 633–637. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 CA203655/CA/NCI NIH HHS/United States
- U10 CA180861/CA/NCI NIH HHS/United States
- R01 CA240910/CA/NCI NIH HHS/United States
- ZIA HL002346/ImNIH/Intramural NIH HHS/United States
- R01 HL103532/HL/NHLBI NIH HHS/United States
- R01 CA216273/CA/NCI NIH HHS/United States
- UG1 CA233338/CA/NCI NIH HHS/United States
- Z01 HL002346/ImNIH/Intramural NIH HHS/United States
- P30 CA006516/CA/NCI NIH HHS/United States
- R01 CA063113/CA/NCI NIH HHS/United States
- P50 CA206963/CA/NCI NIH HHS/United States
- R35 GM122524/GM/NIGMS NIH HHS/United States
- P01 CA206978/CA/NCI NIH HHS/United States
- P01 CA081534/CA/NCI NIH HHS/United States
- R01 DK087992/DK/NIDDK NIH HHS/United States
- R01 CA155010/CA/NCI NIH HHS/United States
- R01 HL116452/HL/NHLBI NIH HHS/United States
- R01 CA173023/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
