

Commensal Microbiota Promote Lung Cancer Development via γδ T Cells
- PMID: 30712876
- PMCID: PMC6691977
- DOI: 10.1016/j.cell.2018.12.040
Commensal Microbiota Promote Lung Cancer Development via γδ T Cells
Abstract
Lung cancer is closely associated with chronic inflammation, but the causes of inflammation and the specific immune mediators have not been fully elucidated. The lung is a mucosal tissue colonized by a diverse bacterial community, and pulmonary infections commonly present in lung cancer patients are linked to clinical outcomes. Here, we provide evidence that local microbiota provoke inflammation associated with lung adenocarcinoma by activating lung-resident γδ T cells. Germ-free or antibiotic-treated mice were significantly protected from lung cancer development induced by Kras mutation and p53 loss. Mechanistically, commensal bacteria stimulated Myd88-dependent IL-1β and IL-23 production from myeloid cells, inducing proliferation and activation of Vγ6+Vδ1+ γδ T cells that produced IL-17 and other effector molecules to promote inflammation and tumor cell proliferation. Our findings clearly link local microbiota-immune crosstalk to lung tumor development and thereby define key cellular and molecular mediators that may serve as effective targets in lung cancer intervention.
Keywords: IL-17; inflammation; lung adenocarcinoma; lung cancer; microbiota; neutrophils; tumor microenvironment; γδ T cells.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Comprehensive Declaration of Interests
The authors wish to list to following outside interests.
T.J. is a member of the Board of Directors of Amgen and Thermo Fisher Scientific and an equity holder in both companies. He is co-Founder and Scientific Advisory Board member of Dragonfly Therapeutics, a co-founder of T2 Biosystems, and a Scientific Advisory Board member of SQZ Biotech; he is an equity holder in all three companies. His laboratory currently receives funding from the Johnson & Johnson Lung Cancer Initiative and Calico. P.C.B. is a consultant to and equity holder in GALT and 10X Genomics, and a consultant to Insitro. S. M. receives funding from the Johnson & Johnson Lung Cancer Initiative. M.M. is a Scientific Advisory Board member for OrigiMed and receives consulting fees for this role. He was a consultant for Foundation Medicine and held equity, some of which was sold to Roche. He receives research support from Bayer and is an inventor of several joint patents and patent applications, none of which have been licensed at the time of this disclosure (12–17–18). In addition, he is an inventor on several patents on EGFR mutations in lung cancer diagnosis, licensed to LabCorp, for which he receives royalties. He is also an inventor on several patent applications on Fusobacterium, none issued or licensed, and the inventor on a patent for pathogen discovery, not licensed, for none of which he receives royalties.
Figures
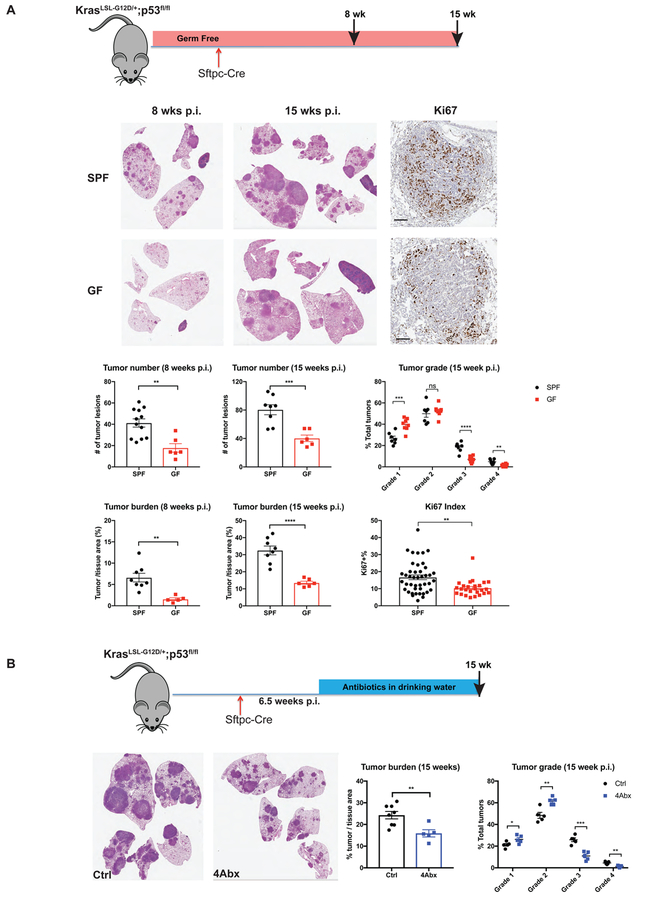
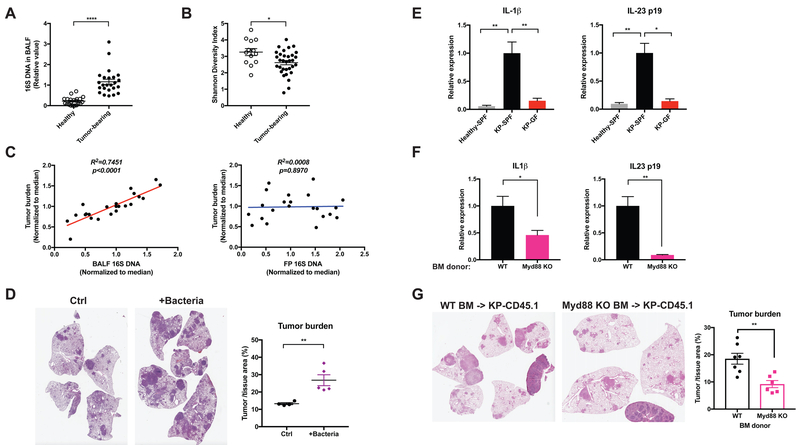
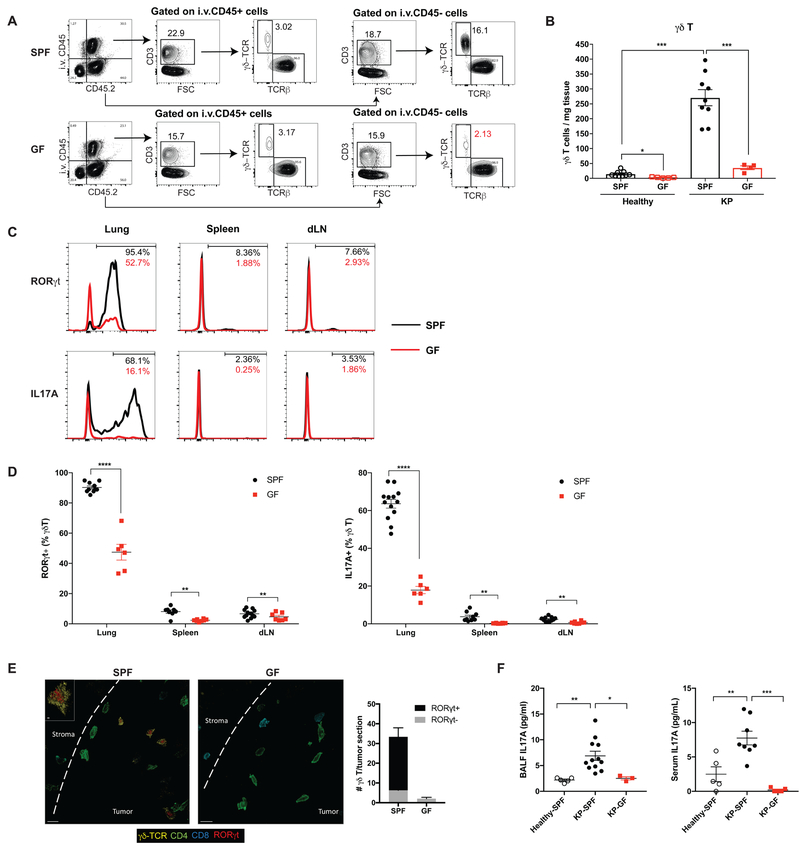
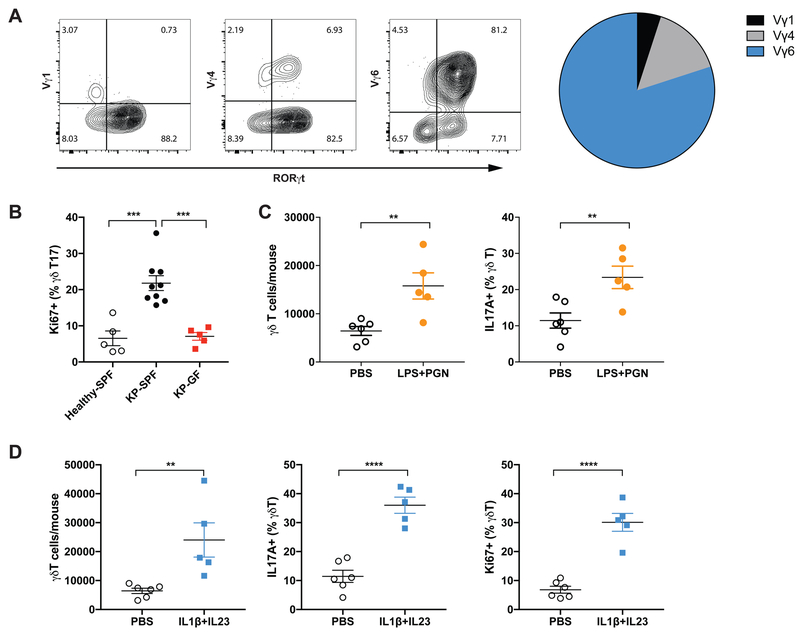
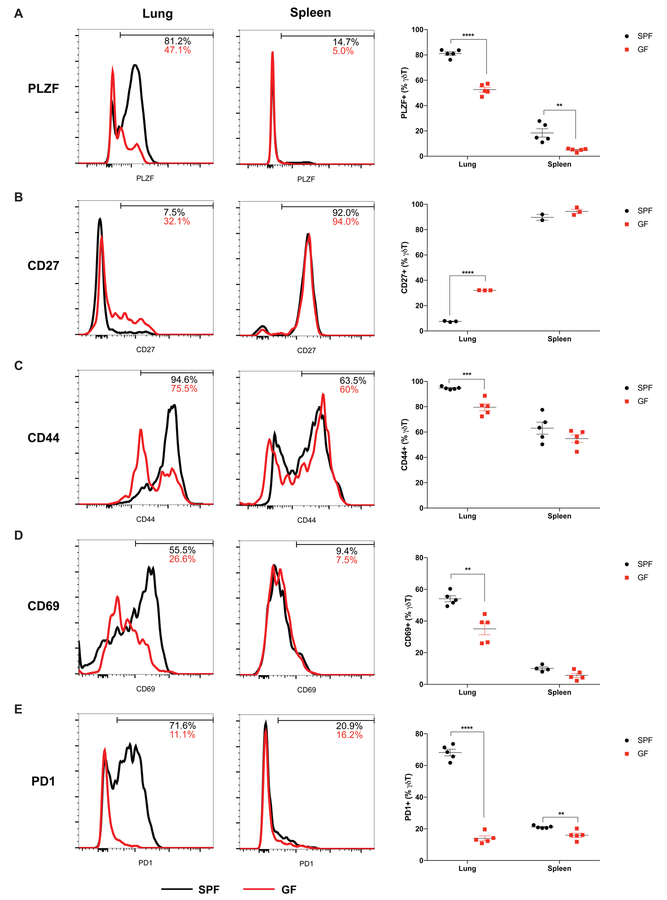
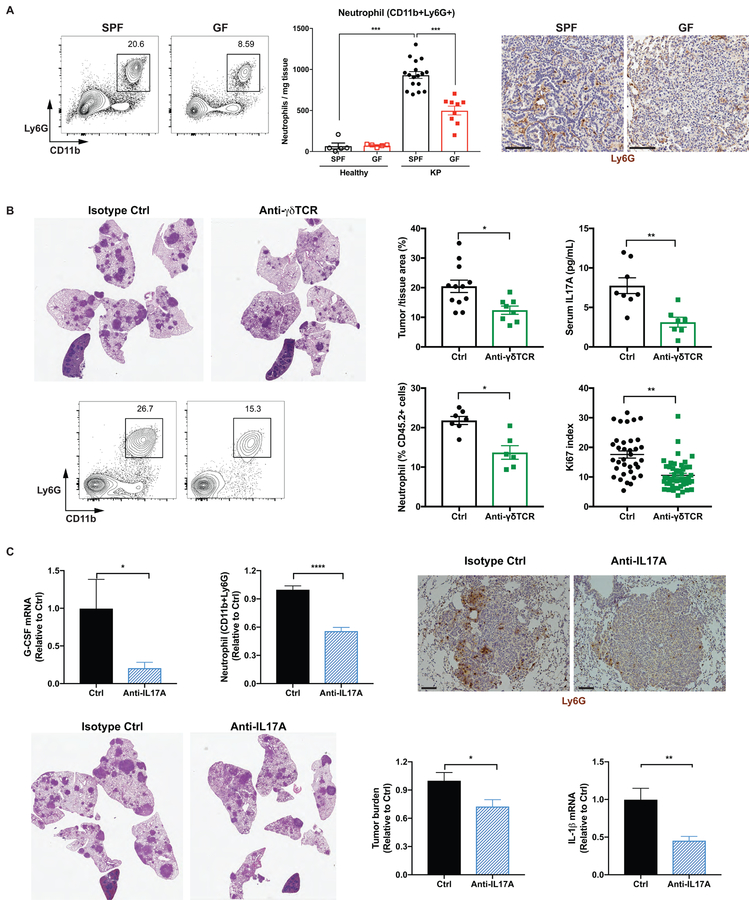
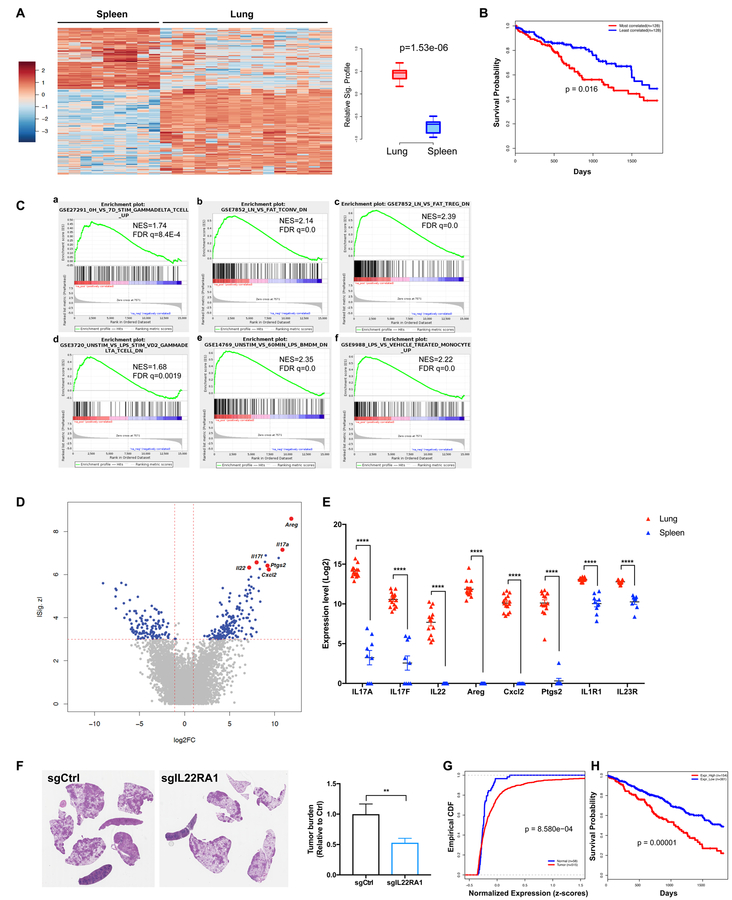
Comment in
-
Lung Microbiota Promote Lung Cancer.Cancer Discov. 2019 Apr;9(4):458. doi: 10.1158/2159-8290.CD-NB2019-019. Epub 2019 Feb 14. Cancer Discov. 2019. PMID: 30765334
-
Microbiota and immune cell interplay.Nat Rev Cancer. 2019 Apr;19(4):182. doi: 10.1038/s41568-019-0122-z. Nat Rev Cancer. 2019. PMID: 30783210 No abstract available.
-
Recent Advances in Lung Immunobiology.Am J Respir Cell Mol Biol. 2019 Dec;61(6):786-788. doi: 10.1165/rcmb.2019-0183RO. Am J Respir Cell Mol Biol. 2019. PMID: 31291124 Free PMC article. No abstract available.
References
-
- Akinosoglou KS, Karkoulias K, and Marangos M (2013). Infectious complications in patients with lung cancer. Eur Rev Med Pharmacol Sci 17, 8–18. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
